
2. Teorias da evolução
Teoria da evolução de Darwin
Durante a viagem de circum-navegação a bordo do navio Beagle (27 de dezembro 1831-2 de outubro de 1836), na dupla função de naturalista e de companheiro de viagem do comandante do navio, o capitão FitzRoy, Darwin, na altura um jovem naturalista em busca de um rumo na carreira e na vida, confrontou-se com duas questões fundamentais que absorviam a atenção dos seus contemporâneos (Pigliucci, 2007):
- Como explicar e qual a história da diversidade biológica que povoa o planeta?
- Por que razão a forma e a função, a flor e a polinização, por exemplo, estão tão estreitamente correlacionadas nos seres vivos?
O afastamento do essencialismo e a aproximação à ideia de evolução, na procura de resposta a estas duas questões fundamentais da biologia, tiveram como ponto de partida o uniformitarismo popularizado pelo geólogo e amigo pessoal de Darwin, Charles Lyell (1797-1875). Lyell é um dos fundadores da geologia moderna e o autor de um importante livro, Principles of Geology, que Darwin leu avidamente durante a viagem do Beagle (Allen, 2014). Para explicar as características geológicas da Terra, Lyell admitiu que estas se deviam à lenta ação de forças ainda hoje atuantes – «o presente é a chave para [entender] o passado», uma ideia simples, mas de grande alcance, cunhada por uniformitarismo (uniformitarianism).
O Beagle deteve-se três anos na América do Sul. No outono de 1835, a cerca de 1.000 km da costa continental sul-americana, nas ilhas do arquipélago dos Galápagos, Darwin deparou-se com várias espécies de aves pertencentes ao género Mimus, de notável semelhança morfológica, quer entre si quer com outras espécies continentais. Regressado a Inglaterra, escreveu em 1836 no seu diário: «Se existe o menor fundamento para essas observações, a zoologia dos arquipélagos será digna de análise; pois tais factos [minariam] a estabilidade das espécies.» Cautelosamente, Darwin considera a hipótese de que espécies tão similares como aquelas descendiam, provavelmente, de um ancestral comum, de origem continental – admite, assim, pela primeira vez que a diversidade biológica não era satisfatoriamente explicada através de um ato único de criação de seres imutáveis (v. Costa, 2009).
Darwin encontrou no seio de cada espécie diferenças morfológicas entre indivíduos. Talvez algumas variações conferissem vantagens na luta por recursos limitados e influíssem no sucesso reprodutivo dos indivíduos que as possuíssem. E a acumulação de características (traits) favoráveis ao longo do tempo poderia explicar a divergência morfológica e o aparecimento de novas espécies. Os fósseis de mamíferos extintos, similares às espécies atuais, que Darwin encontrou nas costas da América do Sul, seriam então elos de uma longa cadeia evolutiva. O seu estudo forneceria não só a evidência da evolução, mas também a possibilidade de reconstruir a história evolutiva dos seres vivos atuais.
O fermento do evolucionismo darwiniano é, porém, anterior à viagem de circum-navegação do Beagle. Entre 1828 e 1831, Darwin foi aluno e usufruiu da amizade de John Henslow (1796-1861), um padre anglicano e um rigoroso investigador da natureza das espécies vegetais e da variação das suas formas. Nas visitas de campo organizadas por este distinto mineralogista e professor de botânica, Darwin aprendeu a colher espécimes botânicos e a observar a variação da forma das plantas. Os seus biógrafos admitem que estas aprendizagens, a par da sua experiência como naturalista na viagem do Beagle (por recomendação de J. Henslow), foram essenciais no processo de aceitação da ideia de evolução e na descoberta do mecanismo da seleção natural (Kohn et al., 2005).
Depois de chegar da viagem do Beagle, Darwin viveu duas outras experiências determinantes no processo de conceção da teoria de evolução por seleção natural: (i) a observação dos métodos de seleção artificial de novas variedades de espécies domésticas, e a (ii) leitura do livro An Essay on the Principle of Population, de Thomas Malthus.
Darwin estudou com minúcia as técnicas usadas por columbófilos e criadores de gado para obter novas raças de animais domésticos, muitas vezes com características exuberantes, através da seleção artificial (artificial selection) da variação biológica intrínseca de cada espécie. Ele próprio dedicou-se a selecionar cruzamentos com raças de pombos domésticos. A analogia entre a seleção artificial de raças domésticas e a seleção natural viria a ser um argumento crucial em A Origem das Espécies (Theunissen, 2012).
Thomas Malthus (1766-1834) foi um clérigo anglicano e demógrafo inglês conhecido por defender que as populações humanas crescem mais rapidamente do que a disponibilidade de recursos. Para Malthus, a escassez era uma inevitabilidade comum a todas as sociedades humanas e a causa eficiente da guerra, da fome e das epidemias. Malthus percebeu que nas sociedades humanas pré-industriais – os efeitos da Revolução Industrial eram ainda incipientes no seu tempo – as taxas de natalidade e de mortalidade tendem a igualar-se e que o rendimento, assim como o número de filhos sobreviventes por mulher, sofreu poucas variações ao longo do tempo (Clark, 2007). Darwin encontrou em Malthus a noção de luta pela sobrevivência (struggle for existence) que adotaria, numa versão menos radical na sua teoria da evolução (Malthus valorizou mais a competição por recursos escassos do que Darwin).
Entre a primeira assunção por escrito de que as espécies mudam, antes referida, e a publicação de A Origem das Espécies decorreram 23 anos. A ideia de evolução era demasiado revolucionária para um homem com uma personalidade temperada, avesso à exposição pública. Darwin começou, finalmente, a escrever A Origem das Espécies em 1856, pressionado por Charles Lyell e pela publicação em 1855 de um artigo por outro importante naturalista britânico, Alfred Russel Wallace (1823-1913), na altura a trabalhar na distante ilha do Bornéu (Figura I.1.3-C). Wallace era um convicto evolucionista – convergiu com todo o mérito, e de forma independente, num mecanismo evolutivo por seleção natural similar ao de Darwin (J. T. Costa, 2009).
A) B)
B) C)
C)
D) E)
E) F)
F)
Figura I.1.3. Seis personagens-chave da história do pensamento evolutivo em biologia. A) Jean-Baptist de Monet (Cavaleiro de Lamarck) (1744-1829), B) Charles Darwin (1809–1882), C) Alfred Russel Wallace (1823-1913), D) August Weismann (1834-1914), E) Ernst Mayr (1904-2005) e F) William Hamilton (1936-2000). [A-E) Wikimedia Commons, F) extraída de Grafen (2004).]
A teoria da evolução por seleção natural foi originalmente proposta, num artigo conjunto de Wallace e de Darwin, em 1858. Ao contrário de Wallace, Darwin conseguiu coligir e publicar um grande número de factos a suportar a teoria da evolução por seleção natural. As 1250 cópias da primeira edição, datada de 1859, de A Origem das Espécies esgotaram imediatamente. O impacto nos meios cultos europeus e norte-americanos foi tremendo. Influentes cientistas, como os botânicos William Hooker (1785-1865) e Asa Gray (1810-1888) ou os zoólogos August Weismann (1834-1914) e Ernst Haeckel (1834-1919), aderiram entusiasticamente às ideias de Darwin. Em contrapartida, os seus detratores não hesitaram em apelidá-lo de «o homem mais perigoso de Inglaterra».
Da teoria da evolução de Darwin, teoria da evolução de Darwin-Wallace ou teoria da evolução por seleção natural extraem-se seis ideias fundamentais (Coyne, 2010):
- A ideia de evolução em si;
- O gradualismo do processo evolutivo;
- A especiação como consequência frequente da evolução;
- A conexão de todos os seres vivos por relações de parentesco (ancestralidade comum);
- A importância fundamental do mecanismo da seleção natural no processo evolutivo;
- A coocorrência da seleção natural com mecanismos não seletivos de mudança evolutiva (menos bem compreendidos por Darwin e entre os quais sobressai a deriva genética).
Kutschera & Niklas (2004) resumem o mecanismo da evolução por seleção natural pugnado por Darwin do seguinte modo:
- Em cada geração nascem mais indivíduos do que aqueles que o meio ambiente pode suportar;
- As características morfológicas e funcionais variam de indivíduo para indivíduo;
- A competição dos indivíduos por recursos escassos gera uma luta pela sobrevivência (struggle for existence) da qual sobrevivem, e atingem a fase reprodutiva, os indivíduos mais aptos (survival of the fittest), com características peculiares de algum modo transmissíveis à descendência;
- As gerações vão-se sucedendo com pequenas modificações estruturais e funcionais;
- Da acumulação de modificações evoluem novas espécies.
Assim concretizado, o mecanismo da evolução por seleção natural processa-se à escala da população, evolvendo três componentes: a) ecológica – os indivíduos competem entre si em determinadas condições ambientais (ambiente seletivo), com contribuições diferenciadas para a geração seguinte (sucesso reprodutivo, fitness); b) genética – parte das características que condicionam a fitness são transmissíveis; c) e de genética populacional – uma população é um arranjo de características transmissíveis (variação genética) com efeito na fitness.
Retorno agora às duas perguntas formuladas no início deste ponto.
Como explicar, então, e qual a história da diversidade biológica que povoa o planeta? Darwin propôs que a especiação e a diversificação dos seres vivos eram consequências diretas da acumulação gradual de pequenas modificações trabalhadas pela seleção natural (Schluter, 2009). E foi mais longe: compreendeu que a ancestralidade comum era a chave para organizar e dar sentido à diversidade biológica, que as relações de parentesco entre as espécies se assemelhavam à ramificação de uma árvore (Figura I.1.4) e que a noção de evolução deveria presidir à construção dos sistemas de classificação. Na Origem das Espécies escreveu: «... o Sistema Natural baseia-se na descendência com modificações ... os caracteres que o naturalista considera revelarem a verdadeira afinidade entre duas ou mais espécies, são os herdados de um ancestral comum, sendo a classificação genealógica ... comunidade de descendência é o vínculo oculto que os naturalistas têm procurado inconscientemente.» (Darwin, 1859).
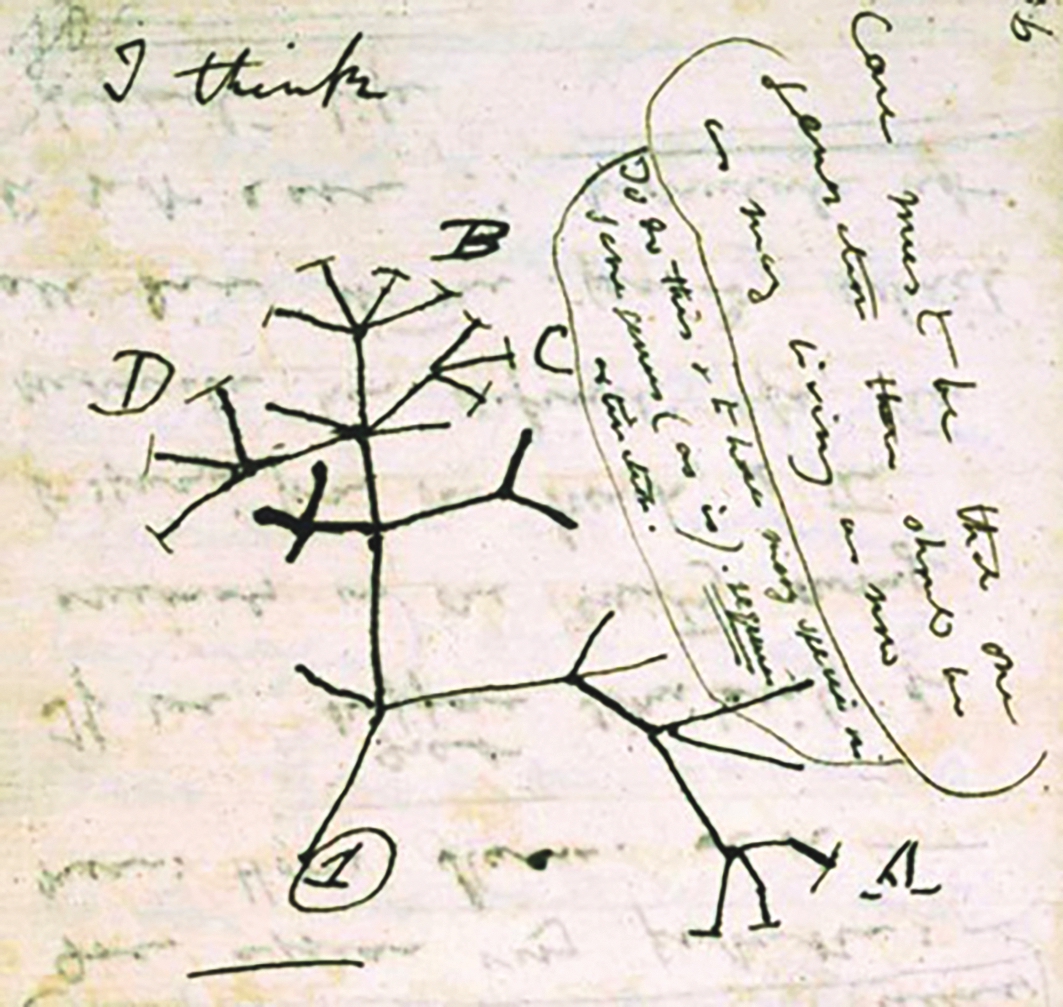
Figura I.1.4. Árvore filogenética idealizada por Charles Darwin num bloco de notas por volta de 1837. N.B. que Darwin reconhece um ancestral comum (1) das espécies agrupadas nos géneros A, B, C e D. [Wikimedia Commons.]
Portanto, a vida na Terra evoluiu de forma gradual a partir de uma espécie-mãe de todas as espécies – hoje em dia entendida como um microambiente povoado por moléculas autorreplicantes –, que evoluiu e se pulverizou em novas linhagens evolutivas (e.g., populações, espécies, clados) (evolutionary lineages), em grande parte (mas não apenas) pela ação da seleção natural. Embora gradual, consoante as linhagens, a evolução desenrola-se a diferentes taxas de evolução (rate of evolution) e tanto mais rapidamente quanto maiores as pressões seletivas. Por exemplo, quando um animal ou uma planta ocupa um novo ambiente, a evolução acelera; à medida que este acumula adaptações ao novo ambiente, a evolução desacelera (v. «Gradualismo filético vs. equilíbrio pontuado»). A divergência de novas espécies – a especiação – segue a mesma lógica: quanto maior a estabilidade ambiental, mais baixa a taxa de aparecimento de novas características e de novas espécies. A explicação da diversidade biológica aperfeiçoou-se, evoluiu, mas a Darwin se deve o «core» da argumentação.
Darwin apercebeu-se, ainda, do papel do isolamento reprodutivo e das pequenas populações na especiação (um embrião conceptual da deriva genética) (Mallet, 2010), e compreendeu que a extinção de taxa de morfologia intermédia explica as descontinuidades morfológicas entre taxa de categoria superior à espécie. «A existência de géneros, famílias, ordens [...] e as suas relações mútuas resultam das extinções que permanentemente ocorrem entre os descendentes divergentes de um stock comum» (Darwin cit. Richards, 2012).
Quanto à segunda questão exposta no início deste capítulo, para Darwin, a ação da seleção natural sobre a variação morfológica e funcional (variação biológica; biologic variation) era suficiente para explicar as correlações forma-função dos organismos. A seleção natural moldou os organismos vivos aos seus nichos ecológicos no passado e continua ativa no presente, porque a evolução é um processo lento e sempre inacabado. Como refere Darwin: «... formas sem fim, as mais belas e as mais maravilhosas, evoluíram e continuam a evoluir» (Darwin, 1859). O mecanismo da seleção natural foi tão perturbador como revolucionário: explica o desenho dos organismos na natureza através de um processo puramente materialístico que não requer nem eventos de criação, nem a intervenção de forças sobrenaturais (J. Coyne, 2010; Mayr, 1982). A Seleção Natural é uma das mais importantes realizações da história do pensamento humano (Dennett, 1995).
Lamarckismo, neolamarckismo e neodarwinismo
A descendência com modificações, i. e., a ideia de evolução e a rejeição do fixismo, foi aceite com alguma rapidez. O papel central no processo evolutivo atribuído por Darwin à seleção natural foi, pelo contrário, recebido com ceticismo. Muitos dos seus contemporâneos consideraram, incorretamente, que a seleção natural eliminava os indivíduos desajustados, mas não explicava a adaptação (Allen, 2014). A agravar esta rejeição, Darwin falhou na explicação das causas da variação e na construção de uma teoria da especiação. Ainda assim, é impressionante que, sem conhecer os mecanismos da variação genética, Darwin tenha sido capaz de intuir que: «Qualquer que seja a causa das pequenas diferenças dos descendentes em relação aos indivíduos parentais – uma causa para cada uma tem de existir –, é a lenta acumulação, por seleção natural, dessas diferenças quando benéficas para os indivíduos que deu origem a todas as mais importantes modificações da estrutura, através das quais os inúmeros seres vivos que povoam a face da Terra são capazes de lutar entre si, e os mais bem-adaptados de sobreviver» (Darwin, 1859).
As reações contra a teoria da evolução por seleção natural recrudesceram no início do século XX, sobretudo em França e na Rússia, com um revivalismo do lamarckismo (v.i.). Este neolamarckismo está associado a um dos episódios mais trágicos da ciência moderna: o assassínio do grande agrónomo russo Nikolai Vavilov (1887-1943). Vavilov, o agrónomo que queria alimentar o mundo, morreu de fome na prisão às mãos de José Estaline, por contestar um charlatão perigoso e opositor acérrimo de Darwin e da genética mendeliana: Trofim D. Lysenko (1898-1976), na altura presidente da Academia de Ciências Agrícolas da URSS (Harlan, 1995).
Darwin não dispunha dos instrumentos conceptuais da genética para aprofundar o mecanismo da seleção natural e para propor uma teoria elaborada e consistente da especiação. Não compreendeu a origem da variação (os conceitos de mutação e recombinação são posteriores) e faltaram-lhe as bases teóricas para associar a especiação a barreiras à troca genética. Essa tarefa, mais o reposicionamento da seleção natural no centro da teoria da evolução, foram realizados pelos proponentes da teoria sintética da evolução (v.i.). Mas, primeiro, houve que rejeitar definitivamente o lamarckismo.
Embora a ideia da evolução organísmica remonte à Grécia Clássica, a primeira teoria coerente da evolução foi proposta na primeira década do século XIX pelo grande naturalista francês Jean-Baptiste de Monet, Cavaleiro de Lamarck (1744-1829), muitos não o sabem, um exímio botânico e o criador da palavra biologia (Figura 3A). A teoria lamarckiana da evolução funda-se, originalmente, em duas supostas leis naturais:
- Lei do uso e do desuso (modificação lamarckiana) – o uso ou desuso causa um gradual fortalecimento, ou enfraquecimento, de uma qualquer característica ao longo do tempo;
- Lei da herança dos caracteres adquiridos – as pequenas alterações geradas pelo uso e desuso são transmitidas e acumuladas de geração em geração.
Para Lamarck, os ancestrais das girafas atuais forçaram o pescoço na tentativa de atingir os rebentos mais elevados das árvores da savana: este uso explica o comprimento inaudito do pescoço das girafas. O não uso da visão atrofiou os olhos de muitos animais cavernícolas. Para explicar o desenvolvimento de novos órgãos nos animais, Lamarck admitiu que novas necessidades geravam fluxos de fluidos no interior do corpo que ao cabo de muitas gerações induziam a diferenciação de novos órgãos (Mayr, 1982), embora falsa, uma explicação revolucionária para a época, porque secundarizava a função de um criador.
O lamarckismo pressupõe que a variação adquirida durante o desenvolvimento de um indivíduo – por causas ambientais ou pelo uso e desuso – pode ser herdada. E. Mayr chamou a este modelo de transmissão de características herança macia (soft inheritance), por oposição à herança forte (hardinheritance), i. e., à transmissão de características estruturais ou fisiológicas geneticamente determinadas cuja génese é independente do meio ambiente. Embora frequentemente omitido nos manuais de biologia, Darwin aceitou a soft e a hard inheritance (Mayr, 1982).
Ainda antes da redescoberta dos trabalhos de genética de G. Mendel, o biólogo alemão August Weismann (Figura 3D) refutou, com uma experiência muito simples, a hipótese da transmissão de caracteres adquiridos (i.e., a soft inheritance) e rejeitou o modelo lamarckiano de evolução. A experiência aparece descrita num texto de 1888 com o título «A Suposta Transmissão de Mutilações»: i) numa primeira etapa, Weismann cortou a cauda de 901 ratos e da sua descendência durante cinco gerações; ii) no final da experiência constatou que os ratos continuaram a desenvolver caudas, sem modificações (Laubichler & Rheinberger, 2006). Uma simples e desagradável experiência destruiu uma aliciante e elegante teoria de grande aceitação. A incorporação das conclusões de Weismann na teoria darwiniana da evolução é conhecida por neodarwinismo (Kutschera & Niklas, 2004).
Weismann expurgou do evolucionismo os elementos lamarckianos ainda tolerados por Darwin e estabeleceu uma distinção fundamental: o papel das células germinativas (transmissoras de informação) face às células somáticas (o corpo). Esta separação — a barreira de Weismann — é válida para os animais, mas não se aplica estritamente às plantas (Lanfear, 2018). Nos animais, a linhagem germinativa é sequestrada precocemente durante a embriogénese: estas células originam os gâmetas, mas não participam na construção dos tecidos do corpo. As plantas, pelo contrário, mantêm células estaminais nos meristemas apicais durante todo o ciclo de vida. Estas células, responsáveis pela formação do corpo vegetativo, podem diferenciar-se posteriormente em estruturas reprodutivas (flores). Consequentemente, as mutações somáticas acumuladas pelas plantas durante o crescimento vegetativo podem incorporar-se na linha germinativa e ser transmitidas à descendência. Embora Weismann reconhecesse a importância da variação e da sua mistura na reprodução sexual, desconhecia ainda a base material da hereditariedade: os genes e os mecanismos de mutação e recombinação.
Teoria sintética da evolução
A biologia contemporânea de Darwin não dispunha de conceitos tão importantes como hereditariedade, gene, alelo, mutação, frequência genética ou deriva genética (Quadro 1). Os fatores hereditários particulados (inherited factors, mais tarde cunhados por genes), com variantes alternativas (alelos), a controlar as características dos organismos, e as leis que regulam a sua transmissão só seriam descobertos após a publicação de A Origem das Espécies,pelo monge católico austríaco Gregor Mendel (1822-1884) (Stenseth et al., 2022). Permaneceriam esquecidos cerca de 35 anos, até à sua redescoberta, em 1900, pelo botânico holandês Hugo de Vries, pelo botânico alemão Carl Correns e pelo agrónomo austríaco E. von Tschermak-Seysenegg (Gliboff, 2015). Darwin não teve conhecimento das descobertas de Mendel, mas Mendel foi fortemente influenciado pela A Origem das Espécies (Fairbanks & Abbott, 2016).
Nas primeiras décadas do século XX, o modelo de transmissão de caracteres qualitativos (mendelianos) – e.g., cor da flor (flores brancas vs. vermelhas) e rugosidade da semente (sementes lisas vs. sementes rugosas) – defendido pelos mendelianos foi considerado incompatível com o gradualismo evolutivo darwiniano. A pergunta de investigação era a seguinte: se a transmissão de características é particulada (descontínua como os átomos) sob a forma de genes (fatores mendelianos, i.e., as unidades básicas e transmissíveis da hereditariedade), como pode a evolução ser contínua? A compatibilização de Darwin e de Mendel foi realizada pelo estatístico inglês Ronald Fisher (1890-1962). Em 1918, Fisher mostrou que os caracteres controlados por um grande número de genes – caracteres quantitativos, e.g., tamanho da folha e forma das folhas – têm uma distribuição contínua; demonstrou ainda que a distribuição das frequências dos valores destes caracteres, ao nível da população, segue uma curva normal e que a sua expressão é fortemente influenciada pelo ambiente (Visscher & Goddard, 2019). De acordo com Fisher, a seleção natural desloca a curva das frequências e a adaptação aumenta ao longo do tempo; a rapidez da deslocação aumenta com a pressão de seleção e a disponibilidade de variação genética na população.
O norte-americano Sewall Wright (1889-1988) compreendeu que as espécies se distribuem espacialmente em pequenas populações que interagem (trocam migrantes) de forma limitada entre si. Wright e o biólogo de origem ucraniana Theodosius Dobzhansky (1900-1975) provaram a importância da deriva genética na evolução das pequenas populações, demonstrando que, em condições de instabilidade ambiental, a seleção incrementa a variação genética (e vice-versa), e que a falta de variação expõe as populações e espécies à extinção (Santillán Zerón, 2009). As conclusões de Wright e de Dobzhansky tiveram um forte impacto no pensamento da época porque contradiziam os defensores da eugenia, os quais, no final do século XIX e primeira metade do século XX, advogavam a noção de raça pura e o desenvolvimento de programas de seleção artificial na espécie humana (entre os quais se contava, aliás, Ronald Fisher).
A «caminhada» científica que culminou na teoria sintética da evolução envolveu outros passos conceptuais e outras descobertas fundamentais, como a teoria cromossómica da hereditariedade de Walter Sutton e Theodor Boveri em 1902 (localização da informação genética nos cromossomas e a explicação celular das leis de Mendel), a distinção entre genótipo (composição genética de um indivíduo) e fenótipo (características estruturais e funcionais de um indivíduo) por Wilhelm Johannsen em 1911, e a teoria da mutação (alterações fortuitas dos genes), bem como a síntese entre hereditariedade, mutação e seleção por Thomas Hunt Morgan e Hermann Joseph Muller, entre outros (Carlson, 2013; Mayr, 1988; Stoltzfus, 2017).
A teoria sintética da evolução (ou síntese evolutiva moderna ou teoria evolutiva padrão), no sentido lato tal qual é geralmente entendida (v.i.), pode ser explicitada do seguinte modo (acompanhar a leitura com os conceitos do Quadro 1 e o esquema da Figura 7) (Futuyma, 2005a; Kingsolver & Pfennig, 2014; Mayr, 1982):
- O indivíduo é a principal unidade de seleção (unit of selection);
- A população é a principal unidade de evolução (unit of evolution);
- A seleção atua nos fenótipos, mas só a seleção de variação fenotípica de base genética tem consequências evolutivas;
- Os indivíduos possuem características estruturais e funcionais influentes na fitness, geneticamente determinadas e transmissíveis;
- As características geneticamente transmissíveis de cada indivíduo são um subconjunto das características disponíveis à escala da população;
- Nova variação genética é produzida continuamente por mutação e recombinação, ou importada por migração;
- As mutações genéticas ocorrem ao acaso (não são direcionadas para uma necessidade adaptativa), cabendo à seleção natural o papel de ordenar essa variabilidade, conferindo direção ao processo evolutivo;
- A seleção natural é a principal força – mas não a única (e.g., deriva genética) – que modela a evolução do fenótipo;
- As características geneticamente transmissíveis positivamente selecionadas têm, por norma, um pequeno efeito no fenótipo dos indivíduos;
- Alterações ambientais alteram as pressões seletivas, favorecendo o aumento da frequência de genótipos que conferem maior aptidão (fitness) nas novas condições;
- A lenta acumulação de novas características transmissíveis resulta numa divergência morfológica e/ou funcional entre populações e, eventualmente, desemboca em especiação;
- A especiação é o processo de formação de populações reprodutivamente isoladas de outras; a Teoria Sintética adota o Conceito Biológico de Espécie (v.i), onde a espécie é definida como um grupo de populações naturais que se cruzam real ou potencialmente e que estão reprodutivamente isoladas de outros grupos;
- Uma parte significativa das aquisições morfológicas e funcionais ocorridas ao longo de milhões de anos de evolução pode ser lida a partir do estudo da biota atual;
- Os mecanismos da macroevolução (diferenciação de taxa de categoria superior à espécie) e da microevolução (diferenciação do ecótipo até à espécie) são comuns.
A teoria sintética da evolução foi desenvolvida para eucariotas sexuados. Em resumo, com ela a evolução passou a ser entendida como a alteração, usualmente gradual, de geração para geração, e à escala da população, da frequência de características geneticamente transmissíveis (genes e alelos), pela ação de mecanismos diversos, sendo a seleção natural ao nível do indivíduo o mais importante, eventualmente culminando em especiação.
Duas das principais consequências desta teoria na biologia foram a rejeição definitiva do lamarckismo e a substituição do pensamento tipológico (= essencialista) lineano por uma nova doutrina metodológica: o pensamento populacional (Mayr, 1982). Sem entrar em grandes detalhes, desvaloriza o papel dos tipos na explicação dos fenómenos populacionais (e.g., extinção, adaptação e especiação), exaltando a importância dos indivíduos per se e da sua variação (Mayr, 1982). A extensão da teoria sintética da evolução às plantas foi consumada pelo botânico norte-americano G. L. Stebbins (Figura 49) no Variation and Evolution in Plants, um livro seminal de 1950.
Os pioneiros da nova síntese valorizaram a recombinação genética em detrimento da mutação na criação de variação genética. Para eles, tanto a micro- como a macroevolução e, implicitamente, a especiação são um produto de alterações das frequências génicas de caracteres quantitativos por seleção natural, sem um envolvimento direto da mutação. E a evolução era intrinsecamente adaptativa. Esta ênfase na genética populacional contrastava, na altura, com os evolucionistas mendelianos como T.H Morgan, defensores de um papel irredutível da mutação.
A descoberta da estrutura em dupla hélice do ácido desoxirribonucleico (DNA) em 1953, por James Watson (1928-2025) e Francis Crick (1916-2004), na sequência de trabalhos prévios de cristalografia da autoria de Rosalind Franklin e de Maurice Wilkins, constitui a objetivação material da genética que nem Darwin nem os seus seguidores neodarwinistas alguma vez anteciparam. O DNA forneceu a base físico-química para a hereditariedade e para as mutações que os arquitetos da síntese neodarwinista haviam postulado de forma largamente teórica. O rápido e extraordinário avanço da biologia molecular expôs a complexidade genómica que, por sua vez, abriu perspetivas inéditas para a Teoria Sintética. Por exemplo, a descoberta de vastas regiões de DNA não codificante, a transferência horizontal de genes, a constatação de que uma grande proporção das mutações ao nível molecular é evolutivamente neutra — fixando-se nas populações por deriva genética e não por seleção natural, como viria a ser formalizado na Teoria Neutralista da Evolução de Motoo Kimura — e, mais recentemente, a revolução epigenética, demonstraram que a arquitetura e a dinâmica da evolução são consideravelmente mais intrincadas do que o modelo adaptacionista de meados do século XX sugeria.
A Teoria Neutralista da Evolução será retomada no capítulo sobre especiação. Na secção «Adaptação vs. aclimatação» apresentam-se os princípios básicos da mecânica da epigenética. Em seguida, relata-se brevemente o impacto da integração da epigenética (com alguns complementos) na Teoria Sintética.
Síntese evolutiva alargada
O processo evolutivo e a origem da biodiversidade têm sido tradicionalmente explicados à luz da síntese evolutiva moderna. Contudo, nas últimas décadas, novas descobertas revelaram mecanismos de hereditariedade, de geração de variação biológica e de especiação que transcendem a genética e a teoria da especiação clássicas, motivando um debate em torno da necessidade de uma Síntese Evolutiva Alargada (Extended Evolutionary Synthesis).
Herança epigenética
A atividade dos genes está sujeita a complexos sistemas de regulação. Entre estes, os mecanismos epigenéticos assumem particular relevância evolutiva, pois é através deles que as condições ambientais influenciam o silenciamento génico e o seu inverso, a desrepressão ou ativação. Estas modificações na expressão dos genes — que não alteram a sequência de DNA — são, pelo menos parcialmente, transmissíveis entre gerações, falando-se, então, de herança epigenética (epigenetic inheritance). Os sistemas de regulação genética sensíveis a fatores ambientais desempenham, aparentemente, um papel na evolução análogo ao das mutações genéticas. Ironicamente, tal sugere a existência de uma componente quase lamarckiana na evolução, onde o ambiente molda diretamente a variação herdável (ainda que de forma mais ou menos temporária).
Evolução reticulada
Nas últimas décadas, tornou-se evidente que a hibridação e processos associados (e.g., alopoliploidia e introgressão), a endossimbiose e a transferência horizontal de genes (THG) desempenham papéis centrais e moldam ativamente a evolução dos seres vivos (Mallet, 2007; Soucy et al., 2015). Quando estes processos de partilha de material genético entre linhagens distintas são relevantes, adota-se o conceito de evolução reticulada (reticulated evolution). Por exemplo, a complexidade celular nos eucariontes está intrinsecamente ligada a eventos endossimbióticos (Lane, 2015). A THG atua como um dos principais motores da diversidade genética e adaptação em procariontes (Doolittle, 1999). Por seu turno, a hibridação/introgressão e a poliploidia são reconhecidamente fundamentais na evolução e na especiação de plantas (Soltis & Soltis, 2009). De realçar que foi datado um brusco aumento da poliploidia nas angiospérmicas em torno da fronteira Cretácico-Paleogénica, coincidindo com um evento maior de extinção global, o que sugere a importância deste mecanismo na sobrevivência e posterior diversificação das linhagens (Vanneste et al., 2014).
Por conseguinte, em diversos grupos taxonómicos, estes mecanismos de fluxo genético desafiam a visão clássica de que a evolução obedece a um padrão de descendência estritamente bifurcado e divergente. Como é realçado no volume III, para representar de forma mais rigorosa a evolução reticulada, o modelo de uma simples "árvore da vida" tem vindo a ser substituído ou complementado por redes ou "teias" filogenéticas (Bapteste et al., 2013).
A evolução reticulada gera sinais filogenéticos contraditórios e dilui a distinção genética e fenotípica entre grupos. Consequentemente, obscurece as relações filogenéticas entre organismos e dificulta os tratamentos taxonómicos (já que as fronteiras morfo-genéticas entre taxa se tornam mais difíceis de estabelecer) (Mallet, 2007).
Plasticidade, evo-devo e construção de nicho
Para além da reticulação e da epigenética, a proposta teórica da síntese evolutiva moderna argumenta que o processo evolutivo é moldado por três outros fatores chave (Laland et al., 2014; Scott-Phillips et al., 2014; West-Eberhard, 2003):
- Biologia evolutiva do desenvolvimento (Evo-Devo): demonstra como pequenas alterações nos genes reguladores do desenvolvimento podem gerar grandes inovações morfológicas; a síntese evolutiva moderna pressupõe que as mutações são absolutamente aleatórias enquanto os biólogos do desenvolvimento alertam para o facto de que alguns caminhos evolutivos (via mutação) são mais prováveis do que outros;
- Plasticidade fenotípica: o ambiente pode induzir variações morfológicas e fisiológicas que antecedem e guiam a própria adaptação genética; por outras palavras, se a alteração do fenótipo em resposta ao ambiente sem alterações genéticas for vantajosa, a seleção natural favorecerá mutações genéticas que, ao longo de gerações, estabilizem esse traço, incorporando-o a posteriori no genoma;
- Teoria da construção de nicho (niche construction theory): o processo pelo qual os organismos modificam o seu ambiente (e.g., a construção de barragens por castores): em vez de evoluírem de forma passiva (como admite a síntese evolutiva moderna) alteram ativamente as pressões seletivas exercidas sobre si próprios e sobre os seus descendentes; assim sendo, a construção de nicho deve ser reconhecida como um processo evolutivo central, em paridade com a seleção natural.
Todos estes casos desafiam a premissa clássica de que a seleção natural é a única força que dá direção à evolução: a canalização evolutiva e a modificação ativa do ambiente também condicionam a trajetória histórica dos organismos.
É necessária uma nova síntese?
A síntese evolutiva moderna consolidou-se em meados do século XX com um foco quase exclusivo na herança vertical, na variação gerada por mutações genéticas aleatórias e graduais, e na genética populacional. Assentava ainda na premissa de um ambiente seletivo independente, face ao qual o organismo é um recetor passivo, e no isolamento reprodutivo (frequentemente alopátrico) como motor primário da especiação. Por conseguinte, a sua formulação clássica apresenta dificuldades teóricas em acomodar a transferência horizontal de genes (THG), a simbiogénese, o surgimento abrupto de novas espécies via poliploidia ou hibridação, e a canalização do processo evolutivo guiada por constrangimentos do desenvolvimento. Ao longo de décadas, a Síntese tendeu a tratar estes mecanismos — bem como a herança epigenética e a construção de nicho — como anomalias raras ou ruído irrelevante na história da vida, algo que a biologia contemporânea demonstrou ser incorreto.
Contudo, a primazia da mutação, da recombinação, da migração, da seleção natural e da deriva genética não está em causa; é meramente complementada por novos mecanismos de herança, de viés no desenvolvimento, na plasticidade e na reticulação. Longe de constituir uma verdadeira «nova síntese» — uma designação talvez presunçosa —, este alargamento concetual não representa um novo paradigma no sentido de uma revolução kuhniana, nem uma rutura com o darwinismo (Pigliucci, 2007). A transmissão de caracteres adquiridos por influência ambiental (soft inheritance), a transferência horizontal, a hibridação ou a construção do nicho não invalidam o papel absolutamente chave da herança genética forte e da seleção natural no processo evolutivo.
Da mesma forma que Darwin identificou a seleção natural sem conhecimentos de genética, os líderes da nova síntese expandiram a teoria darwiniana da evolução sem suspeitarem dos intrincados mecanismos de controlo molecular da vida. A biologia moderna, entretanto, veio conectar o processo evolutivo a todos os níveis de complexidade biológica. A síntese evolutiva moderna mantém-se, ainda assim, como o referencial de cúpula porque foi capaz de explicar o essencial do processo evolutivo à escala da população — a dimensão fundamental para analisar o funcionamento e a organização da diversidade da vida.