
1. Conceito de planta
No século IV a.C., Aristóteles dividiu os seres vivos em dois grandes grupos, depois de Carl Linnaeus (1707-1778) categorizados ao nível do reino: os reinos Plantae e Animalia (= Metazoa). Em 1866, o zoólogo alemão Ernst H. Haeckel (1834-1919) reconheceu que nem todos os seres vivos são animais ou plantas e sugeriu, então sem grande sucesso no meio académico, a criação de um novo reino – o reino Protista – para absorver os atuais procariotas, os protozoários, as algas e os fungos, um imenso biota entretanto desvendado pelos progressos da microscopia. Durante boa parte do século XX, os livros-texto de botânica, além das plantas terrestres, incluíram no reino das Plantas todo o tipo de algas, os fungos e até alguns grupos de bactérias (Hagen, 2012). Somente em 1962, quase cem anos depois da proposta de E. Haeckel, os microbiologistas R. Y. Stanier e C. B. van Niel clarificaram a dicotomia fundamental da vida entre procariotas e eucariotas, identificada na década de 1920 pelo biólogo francês Édouard Chatton (1883-1947), eliminando, em definitivo, a clássica oposição planta-animal.
Robert Whittaker (1920-1980) propôs, com um enorme êxito, um sistema de classificação alternativo à aproximação lineana, com cinco reinos (Whittaker, 1969): Monera, Protista, Animalia, Fungi e Plantae. Este conhecido sistema tem uma índole funcional e ecológica – R. Whittaker era um ecólogo de vegetação.
O sistema de classificação de Whittaker baseia-se em três critérios (Hagen, 2012):
- Nível de organização – procariotas vs. eucariotas; unicelularidade vs. multicelularidade;
- Modo de nutrição – autotrofia, ingestão ou absorção;
- Nível trófico – produtores, consumidores ou decompositores.
As plantas, sensu Whittaker, são seres eucariotas, pluricelulares, autotróficos e produtores.
Este sistema não expressa qualquer tipo de relação evolutiva (= relação filogenética). Por exemplo, para Whittaker uma alface-do-mar (Ulva lactuca) é uma planta; outra alga verde fotossintética unicelular evolutivamente próxima da alface-do-mar, um protista. Os conhecimentos de bioquímica, fisiologia, genética e biologia evolutiva acumulados nas últimas décadas, a par da progressiva aceitação do princípio da monofilia (principle of monophyly) – os taxa têm de incluir todos, e apenas, os descendentes de um ancestral comum (volume II) –, implicaram uma redução significativa da circunscrição do conceito de planta e a substituição do sistema ecológico/funcional de R. Whittaker por sistemas de classificação filogenéticos. Afinal nem todas as plantas são pluricelulares e nem todos os seres multicelulares autotróficos são plantas.
As plantas são eucariotas (domínio Eukaryota), um dos três domínios (domains) da vida celular[1] definidos por Woese et al. (1990). Embora a origem dos eucariotas permaneça um dos maiores desafios da biologia evolutiva atual (Lane, 2015), o conhecimento das relações filogenéticas entre os grandes grupos de eucariotas está a evoluir rapidamente e a ser incorporado nos modernos sistemas de classificação.
Apesar das plantas, dos animais e dos fungos agruparem a vasta maioria das espécies de eucariotas conhecidas, representam uma pequena porção da diversidade eucariótica. O sistema de classificação de Adl et al. (2012) reconhece cinco supergrupos[2] de eucariotas: Opisthokonta, Amoebozoa, Excavata, SAR[3] e Archaeplastida. Grosso modo, os Opisthokonta incluem, entre outros seres, os fungos (Fungi) e os animais (Metazoa); as Amoebozoa e as Excavata reúnem essencialmente seres unicelulares; os oomicetas (Peronosporomycetes, = Oomycetes)[4], as algas castanhas (Phaeophyceae) e as diatomáceas (Diatomea, = Bacillariophyta) são os grupos de SAR mais conhecidos; as plantas são colocadas no supergrupo Archaeplastida. No esquema de Burki et al. (2020) admitem-se sete supergrupos e diversos clados de parentescos não resolvidos; as Archaeplastida são o único supergrupo considerado por Adl et al. (2012) de circunscrição conservada. Aguardam-se severas alterações nas Archaeplastida com a descoberta de dois pequenos grupos de protozoários não fotossintéticos aparentados com as algas vermelhas, Rhodelphida e Picozoa (Gawryluk et al., 2019; Schön et al., 2021).
Para K. Niklas todos os eucariotas fotossintéticos são plantas (Niklas, 2000). Outros autores restringem o conceito às plantas terrestres (Plantae s.str.); a maioria opta por uma solução intermédia. Por conseguinte, o nome Plantae é equívoco porque, desde a fundação da moderna nomenclatura biológica por Linnaeus, nos meados do século XVIII, foi usado com demasiados sentidos. Uma solução parcimoniosa corrente para este inconveniente passa pela substituição do nome por Archaeplastida, com a categoria não formal de supergrupo (Adl et al., 2012), agrupando todos os descendentes do evento de endossimbiose primária do cloroplasto.
A figura que se segue resume as relações filogenéticas e a nomenclatura dos grandes grupos de arqueoplastidas (plantas), por inerência os mais basais (antigos), com base na bibliografia mais recente (Figura I.1.1) [5],[6].
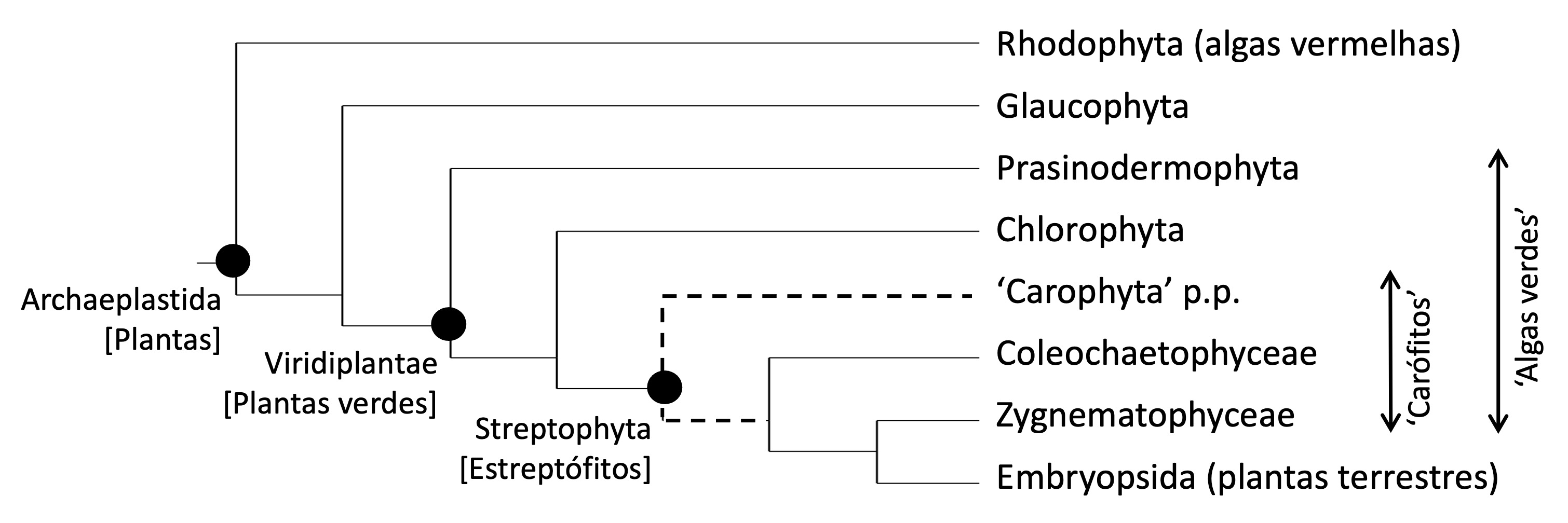
FIGURA I.1.1. Relações evolutivas entre os grupos basais de plantas. N.b.: as algas verdes são um grupo parafilético porque não incluem todos os descendentes de um ancestral comum (excluem as plantas terrestres) estando, por isso, grafados entre aspas simples. Os grupos de 'carófitos' estão parcialmente omitidos. Depreende-se da figura que as plantas terrestes são estreptófitos terrestrealizados com um ancestral algal. [Filogenia baseada em Bowles et al. (2023) e Bierenbroodspot et al. (2024).]
As arqueoplastidas incorporam os seguintes grupos monofiléticos (= clado, monphyletic group, clade), i.e., que incluem todos, e apenas, os descendentes de um ancestral comum:
- Arqueoplastidas = algas vermelhas + glaucófitos + plantas verdes.
Os três grupos que constituem as arqueoplastidas possuem cloroplastos cuja origem remonta a uma cianobactéria fagocitada por um eucariota ancestral heterotrófico (já portador de mitocôndrias) num evento único de endossimbiose (endossimbiose primária). Os plastídios fotossintéticos dos restantes grupos de seres autotróficos (e.g. algas castanhas) foram adquiridos pela endossimbiose de algas vermelhas ou verdes em diferentes hospedeiros eucarióticos (endossimbioses secundárias). Em raros casos ocorreram endossimbioses terciárias ou substituições secundárias de plastídios.
As plantas verdes, o maior dos grupos de arqueoplastidas, repartem-se por três grandes linhagens:
- Plantas verdes = prasinodermófitos + clorófitos + estreptófitos.
As chamadas ‘algas verdes’ englobam prasinodermófitos, clorófitos e parte dos estreptófitos. As aspas simples indicam que as ‘algas verdes’ são um grupo parafilético (= grado, paraphyletic group, grade), i.e., que não inclui todos os descendentes do ancestral comum das plantas verdes. Com efeito, os estreptófitos são constituídos por um grupo parafilético de ‘algas verdes’ longamente explorado no volume II – as ‘Charophyta’ (= ‘carófitos’ e ‘algas carófitas’) –, mais as plantas terrestres, seus descendentes diretos.
Então:
- Estreptófitos = ‘carófitos’ + plantas terrestres.
Por fim, as plantas terrestres abrangem quatro grandes grupos monofiléticos (Figura I.1.2) (Figura I.1.3) (Leebens-Mack et al., 2019):
- Plantas terrestres = briófitos + licopodiófitos[7] + fetos + plantas com semente.
Sendo:
- Plantas vasculares = licopodiófitos + fetos + plantas com semente;
- Briófitos = antóceros + hepáticas + musgos;
- Fetos = equisetófitos + ophioglossófitos + marattiófitos + polipodiófitos;
- Plantas com semente = gimnospérmicas + angiospérmicas;
- Gimnospérmicas = Ginkgoidae + Cycadidae + Cupressidae + Pinidae + Gnetidae;
- Angiospérmicas (= plantas com flor) – clados mais importantes discutidos no volume III.
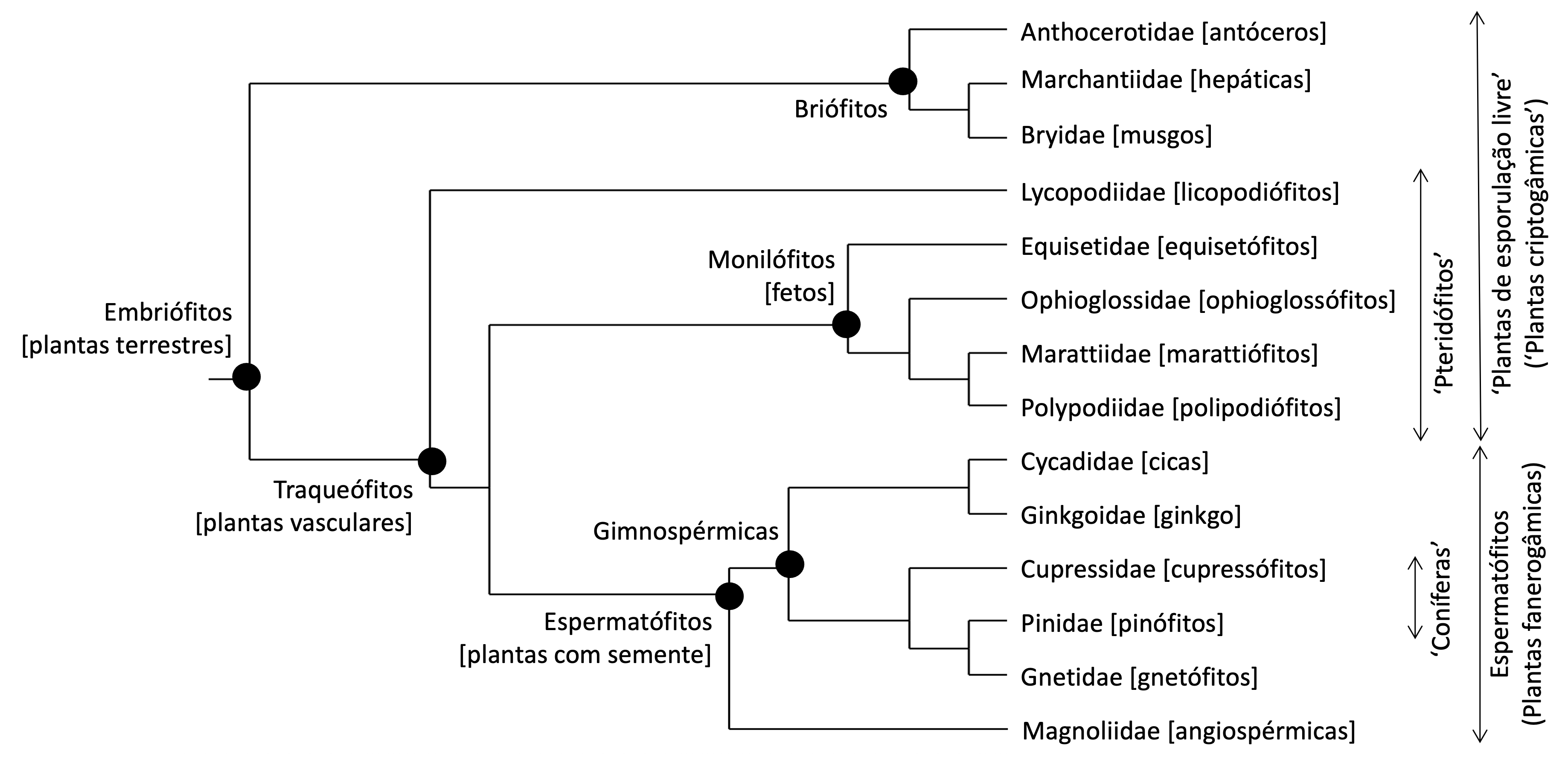

FIGURA I.1.2. Relações evolutivas entre os grandes grupos atuais de plantas terrestres. A) Filogenia baseada em Bechteler et al. (2023) para os briófitos, Testo & Sundue (2016) e Schuettpelz et al. (2016) para os fetos, Ran et al. (2018) e Yang et al. (2022) para as gimnospérmicas e Zuntini et al. (2024) para as angiospérmicas; nomenclatura resumida no Quadro 1.1. B) Conocephalum conicum (Conocephalaceae, Marchantiidae). C) Bryum dichotomum (Bryaceae, Bryidae). D) Phaeocerus sp. (Notothyladaceae, Anthocerotidae). E) Huperzia dentata (Huperziaceae, Lycopodiidae). F) Equisetum telmatea (Equisetaceae, Equisetidae). G) Botrychium lunaria (Ophioglossaceae, Ophioglossidae). H) Marattia laevis (Marattiaceae, Marattiidae). I) Culcita macrocarpa (Culcitaceae, Polypodiidae). J) Cycas revoluta (Cycadaceae, Cydadidae). K) Ginkgo biloba (Ginkgoaceae, Ginkgoidae). L) Metasequoia glyptostroboides (Cupressaceae, Cupressidae). M) Larix decidua (Pinaceae, Pinidae). N) Ephedra fragilis (Ephedraceae, Gnetidae). O) Agapanthus africanus (Amaryllidaceae, Magnoliidae). P) Lophira lanceolata (Ochnaceae, Magnoliidae). [A) original; B-F e H-P) fotografias do autor; G) Wikimedia Commons.]
Os briófitos são, muito provavelmente, monofiléticos (Bechteler et al., 2023; Harris et al., 2020). Como se verá no volume II, as primeiras plantas a colonizarem a terra firma, algures no final do Câmbrico (541-485 M.a.) ou no início do Ordovícico (485-458 M.a.), pertenciam a um grupo, hoje extinto, provavelmente taloso, com um corpo gametofítico semelhante ao dos atuais antóceros e de muitas hepáticas (Edwards et al., 2014; Morris et al., 2018). São igualmente monofiléticos três importantes grupos sem categoria formal: os traqueófitos, os fetos e as plantas com semente.
Os ‘pteridófitos’ agrupam licopodiófitos e fetos propriamente ditos. Apesar de frequente na bibliografia, o uso do termo “pteridófito” deve ser descontinuado, como está a acontecer com os termos criptogâmico e fanerogâmico. Os briófitos, os licopodiófitos e os fetos são ‘plantas de esporulação livre’ (free sporing plants), i.e., disseminam-se por esporos. Nas plantas com semente, os esporos ♀ estão retidos no interior do primórdio seminal e em sua vez dispersam-se sementes.
As plantas com semente são o objeto deste conjunto de livros.
No Quadro I.1.1. faz-se um enquadramento taxonómico dos principais grupos de plantas terrestres (relações evolutivas explicitadas nas Figuras I.1.1 e I.1.2).
|
Grandes grupos de plantas abordados neste texto. Baseado em Bowles et al. (2023), Chase & Reveal (2009), Christenhusz et al. (2011) e Schuettpelz et al. (2016). |
|||
|
Categoria taxonómica |
Grupo taxonómico |
Nome vulgar (português) |
Nome vulgar (inglês) |
|
Ordenação hierárquica dos grandes grupos de plantas terrestes |
|||
|
Sem categoria formal |
Archaeplastida (= Plantae s.l., Primoplantaae) |
Arqueoplastidas, plantas |
Plants |
|
Sem categoria formal |
Viridiplantae* |
Plantas verdes |
Green plants |
|
Sem categoria formal |
Streptophyta |
Estreptófitos |
Streptophytes |
|
Classe |
Embryopsida** |
Plantas terrestres, embriófitos |
Terrestrial plants, embryophytes |
|
Sem categoria formal |
Bryophyta |
Briófitos |
Bryophytes |
|
Subclasse |
Anthocerotidae |
Antóceros |
Hornworts |
|
Subclasse |
Marchantiidae |
Hepáticas |
Hepatics, liverworts |
|
Subclasse |
Bryidae |
Musgos |
Mosses |
|
Sem categoria formal |
Tracheophyta |
Plantas vasculares, traqueófitos |
Vascular plants |
|
Subclasse |
Lycopodiidae |
Licófitos, licopodiófitos, licopódios |
Lycophytes, polydiophytes, lycopods |
|
Sem categoria formal |
Monilophyta |
Fetos, monilófitos |
Ferns, monilophytes |
|
Subclasse |
Equisetidae |
Equisetidas, equisetófitos, equisetas, cavalinhas |
Horsetails |
|
Subclasse |
Ophioglossidae*** |
Ophioglossidas, ophioglossófitos, psilotófitos**** |
Ophioglossophytes |
|
Subclasse |
Marattiidae |
Marattiidas, marattiófitos |
Marattiophytes |
|
Subclasse |
Polypodiidae |
Polipodiidas, polipodiófitos, fetos verdadeiros, fetos leptoesporangiados |
True ferns, leptosporangiate ferns |
|
Sem categoria formal |
Spermatophyta |
Plantas com semente, espermatófitos |
Seed plants, spermatophytes |
|
Subclasse |
Ginkgoidae |
Ginkgoídas, ginkgófitos, ginkgos |
Ginkgophytes |
|
Subclasse |
Cycadidae |
Cicadidas, cicadófitos, cicas |
Cycadophytes |
|
Subclasse |
Cupressidae***** |
Cupressidas, cupressófitos |
Cupressophytes |
|
Subclasse |
Pinidae |
Pinidas, pinófitos |
Pinophytes |
|
Subclasse |
Gnetidae |
Gnetidas, gnetófitos |
Gnetophytes |
|
Subclasse |
Magnoliidae (= Angiospermae) |
Angiospérmicas, magnoliófitos, magnoliidas, plantas com flor |
Flowering plants, angiosperms |
|
Outros grandes grupos de plantas usados neste texto (ordenação não hierárquica) |
|||
|
Sem categoria formal |
Rhodophyta |
Algas vermelhas; rodófitos |
Rhodophytes, red algae |
|
Sem categoria formal |
Prasinodermophyta |
Prasinodermófitos |
Prasinodermophytes |
|
Sem categoria formal |
Glaucophyta |
Glaucófitos |
Glaucophytes |
|
Sem categoria formal |
Chlorophyta |
Clorófitos |
Chlorophytes |
|
Sem categoria formal |
‘Charophyta’ |
‘Carófitos’ |
Charophytes |
|
Sem categoria formal |
Gymnospermae |
Gimnospérmicas |
Gymnosperms |
|
* O nome Viridiplantae é mais antigo pelo que não deve ser rejeitado em favor de Chloroplastida. Outros nomes citados na bibliografia: Chlorobionta, Chlorobiota ou Chloroplastida. ** Nome correto para a classe que expressa a retenção do embrião diploide no arquegónio (Pirani & Prado, 2012). Outros nomes científicos disponíveis: Equisetopsida, Embryobionta, Embryophyta e Archegoniatae. *** Psilotidae (psilotófitos) até há bem pouco tempo. ***** Designação mais corrente na bibliografia. **** Pinidae s.l. (Pinales + Araucariales + Cupressales) é parafilética (Leebens-Mack et al., 2019; Ran et al., 2018) . A solução passa por recuperar a subclasse Cupressidae (Araucariales + Cupressales) não admitida por Christenhusz et al. (2011). |
|||
Uma vez que este texto versa as plantas com semente, e as ‘plantas de esporulação livre’ são ciclicamente recordadas, impõe-se, desde já, uma importante questão: o que é uma planta terrestre? As plantas terrestres são um taxon monofilético de plantas adaptadas a ambientes terrestres, secundariamente aquáticas, cujos elementos partilham um conjunto alargado de características funcionais e estruturais herdado do ancestral comum, resumidas no Quadro 1.2. No Quadro 1.3 comparam-se vários grupos de plantas, grupos algais incluídos.
|
QUADRO 1.2. Principais características estruturais e funcionais das plantas terrestres. Baseado em Ingrouille & Eddie (2006) com adições. |
|
|
Característica |
Descrição |
|
Estrutura celular eucariótica |
Células com um núcleo e outras estruturas intracelulares complexas encerradas por membranas. |
|
Multicelularidade |
Indivíduos com mais de uma célula, células estas com diferentes funções e interdependentes entre si. |
|
Imobilidade |
Organismos fixos ao substrato durante grande parte do seu ciclo biológico, com fases de dispersão breves sob a forma de propágulos (e.g., rizomas), esporos (nas ‘plantas de esporulação livre’), ou pólen e sementes (nos espermatófitos). |
|
Estrutura modular |
Indivíduos constituídos pela repetição de unidades multicelulares discretas, i.e., por módulos, de grande autonomia funcional (semiautónomos) (v. «Estrutura modular. Totipotência celular. Plasticidade»). |
|
Elevada plasticidade fenotípica |
Capacidade de um determinado genótipo alterar a sua morfologia em função das condições ambientais (v. «Estrutura modular. Totipotência celular. Plasticidade»). |
|
Crescimento indeterminado |
Indivíduos crescem continuamente até à senescência/morte, ainda que este crescimento possa ser interrompido por períodos de quiescência/dormência mais ou menos prolongados (v. «Organização do corpo das plantas com semente»). |
|
Paredes celulares celulósicas |
Estrutura rígida que envolve o protoplasma maioritariamente constituída por celulose, um polímero de α-glicose. |
|
Cutícula |
Camada externa não celular, cerosa, de proteção dos órgãos aéreos primários (v. «Epiderme»). |
|
Revestimento de algumas estruturas reprodutivas com esporopolenina |
Biopolímero complexo, de composição química mal conhecida, muito resistente à agressão química, empregado pelas plantas na proteção de esporos e grãos de pólen contra os raios UV e a dessecação (v. «Pólen»). |
|
Fotoautotrofia |
Produzem compostos orgânicos complexos e ricos em energia a partir de moléculas inorgânicas simples (e.g., H2O, CO2, K+, e NO3-), e da energia química (sob a forma de ATP) e do poder redutor (sob a forma de NADPH2) gerados pelas moléculas de clorofila excitadas pela luz solar. |
|
Órgãos assimiladores* com uma elevada relação superfície/volume |
Elevada relação superfície/volume conseguida através da redução da espessura das folhas e do diâmetro das raízes; uma consequência da fototrofia e do consumo de alimentos inorgânicos diluídos (v. «Volume e superfície nas plantas»). |
|
Estomas |
Pequenas aberturas com células-guarda por onde se processam as trocas gasosas com o exterior na superfície dos órgãos aéreos primários («Anatomia da folha [nomofilo]»). Os estomas são exclusivos do esporófito; não têm estomas os esporófitos das hepáticas e de alguns grupos de musgos e antóceros (Merced & Renzaglia, 2017). |
|
Órgãos especializados na absorção de nutrientes do solo |
Rizoides (nos briófitos e no protalo dos ‘pteridófitos’) ou sistema radicular (no esporófito dos ‘pteridófitos’ e das plantas com semente) (v. «Natureza e funções da raiz»). |
|
Órgãos fotossintetizadores suportados por um sistema tubular rígido |
Sistemas de caules com folhas s.l., i.e., com filídios nos musgos, microfilos nos licopodiófitos ou megafilos nos eufilófitos (= fetos + plantas com semente) (v. «Natureza e funções da folha»). |
|
Ciclo de vida haplodiplonte heteromórfico |
Com meiose desfasada da fecundação e alternância de duas gerações – fases haploide (gametófito) e diploide (esporófito) – de distinta morfologia (v. «Ciclos de vida das plantas terrestres»). |
|
Anterídios e arquegónios |
Órgãos onde se diferenciam, respetivamente, gâmetas ♂ e ♀ (tremendamente simplificados nos espermatófitos; «v. Ciclos de vida das plantas terrestres»). |
|
Esporângios |
Órgãos onde se diferenciam esporos («Ciclos de vida das plantas terrestres»). |
|
Embrião |
Rudimento do esporófito; estrutura multicelular protegida por um tecido multicelular haploide (nos ‘briófitos’, ‘pteridófitos’ e gimnospérmicas) ou triploide (nas angiospérmicas) (v. «Ciclos de vida das plantas terrestres»). |
|
* Entende-se por assimilação a incorporação e conversão de nutrientes no protoplasma, processo que nas plantas envolve a fotossíntese nos órgãos herbáceos aéreos (caules primários e folhas) e a absorção de nutrientes pelas raízes. |
|
|
QUADRO 1.3 Comparação resumida dos grandes grupos de plantas |
|||||||||
|
Grupo |
N.º de |
Geração |
Embrião |
Esporulação livre |
Estomas no esporófito |
Tecidos vasculares |
Cormo |
Semente |
Flor e fruto |
|
Algas glaucófitos algas vermelhas algas verdes |
>21 000 15 7000 14 000 |
Sem alternância de gerações e domínio do gametófito** |
Não |
Sem esporos |
Não |
Não |
Não |
Não |
Não |
|
‘Briófitos’ hepáticas musgos antóceros |
>13 000 7266 16 000 220 |
Gametófito (n) |
Sim |
Sim |
Não Sim** Sim** |
Não |
Não |
Não |
Não |
|
Licófitos |
1340 |
Esporófito (2n) |
Sim |
Sim |
Sim |
Sim |
Não |
Não |
Não |
|
Fetos ophioglossófitos marattiófitos polipodiófitos |
10 892 110 120 10 662 |
Esporófito (2n) |
Sim |
Sim |
Sim |
Sim |
Sim |
Não |
Não |
|
Gimnospérmicas cicadófitos ginkgófitos cupressófitos pinófitos gnetófitos |
1045 317 1 390 225 112 |
Esporófito (2n)
|
Sim |
Não |
Sim |
Sim |
Sim |
Sim |
Não |
|
Angiospérmicas |
246 366 |
Esporófito (2n) |
Sim |
Não |
Sim |
Sim |
Sim |
Sim |
Sim |
|
* Estimativa do número de espécies extraída de Roskov et al. (2019). ** Com exceções. |
|||||||||
[1] Por definição, excluindo vírus e priões.
[2] Categoria não formal, i.e., não reconhecida pelo Code of Nomenclature for Algae, Fungi, and Plants (ICN) (Turland et al., 2018).
[3] Acrónimo de Stramenopiles, Alveolata e Rhizaria.
[4] Os oomicetas, como os fungos, produzem micélio e esporos. Incluem os conhecidos míldios da videira‑europeia (Plasmopara viticola) e da batateira (Phytophthora infestans). Foi demonstrado que não estão evolutivamente relacionados com os fungos e que a partilha de uma morfologia comum é um caso de convergência evolutiva.
[5] Reparar que não se categorizam formalmente Eukaryota, Archaeplastida, Viridiplantae, Streptophyta e Chlorophyta.
[6] Os nomes comuns em inglês estão escritos em itálico e inscritos entre parêntesis.
[7] Mais correto licopodiófitos do que licófitos (Christenhusz et al., 2018).