
1. A célula vegetal
Teoria celular
A célula foi descrita pela primeira vez pelo inglês Robert Hooke (1635-1703), em 1665, a partir de cortes finos de cortiça. Ao holandês Antonie van Leeuwenhoek (1632–1723) cabe a honra da primeira observação de células vivas e de microrganismos unicelulares. As evidências observacionais subsequentemente acumuladas, assistidas pelos progressos da microscopia ótica, culminaram século e meio depois na teoria celular. Esta teoria fundamental da biologia, originalmente proposta pelo botânico Matthias Schleiden (1804-1881) e pelo fisiologista Theodor Schwann (1810-1882), ambos de nacionalidade alemã, em 1839, postula que: (i) todos os seres vivos são constituídos por células; (ii) a célula é o elemento fundamental da organização da vida; (iii) todas as células descendem de outra célula (Mazzarello, 1999).
A vida, tal como a conhecemos, evoluiu uma única vez – todos os seres vivos têm, em última instância, um ancestral comum, cujas características quedam por clarificar (Lane, 2015). A natureza celular do primeiro ser vivo é, porém, inequívoca – a célula é uma plesiomorfia, i.e., um carácter ancestral, neste caso de toda a vida terrestre. A estrutura da célula vegetal foi razoavelmente conservada pela evolução, i.e., varia pouco de taxon para taxon e, por isso, o estudo aprofundado da citologia vegetal tem em muitas formações académicas um interesse marginal.
Parede celular
O LECA (Last Eukaryotic Common Ancestor), o ancestral de todos os eucariotas atuais, era heterotrófico. Provavelmente, alimentava-se de procariotas ingeridos por fagocitose (Martin et al., 2017). Um evento incompleto de fagocitose de uma cianobactéria ocorrido há cerca de 2.100 M.a. marca o nascimento das arqueoplastidas. O endossimbionte cianobacteriano entretanto evoluiu num plastídeo fotossintético e os componentes orgânicos da célula vegetal passaram a ser produzidos no seu interior, a partir dos esqueletos carbonados fornecidos pela fotossíntese (Gibson et al., 2017; Sánchez-Baracaldo et al., 2017). A célula vegetal pôde, então, especializar-se no consumo de substâncias minerais simples, capazes de penetrar na célula por absorção – e a fagocitose volveu inútil[1]. A absorção, ao contrário da fagocitose, é compatível com o desenvolvimento de uma parede celular. O agravamento dos riscos de rutura das membranas celulares vegetais em ambientes hipotónicos resultante da concentração no citoplasma de compostos orgânicos solúveis provenientes da fotossíntese criou um ambiente seletivo propício para a evolução da parede celular (Sarkar et al., 2009). A parede celular é também uma óbvia estrutura de resistência contra predadores e parasitas. A autotrofia está, portanto, à cabeça da longa cadeia causal que explica o aparecimento de uma parede rígida nas arqueoplastidas. Mais tarde, essa parede celular foi determinante na proteção contra a radiação UV, na rigidez estrutural e na contenção das perdas de água necessárias para a conquista da terra emersa (Volume II).
A parede celular primária das células somáticas (= não reprodutoras) das plantas vasculares diferencia-se ainda no interior do meristema. A parede celular primária é uma armadura resistente, e ao mesmo tempo flexível e porosa, de modo a permitir o alongamento celular e o movimento de água e outras substâncias nos espaços intercelulares. É constituída por três tipos de polissacarídeos (Refrégier et al., 2012): (i) microfibrilas de celulose (um polímero hidrófilo de β-D-glucose), (ii) entrelaçadas por hemiceluloses (polímeros de xiloglucano), (iii) numa matriz de pectinas (um polímero de ácido galacturónico). A celulose representa apenas cerca de 15% do peso seco da parede primária. As células alongam-se em consequência da pressão exercida pelo protoplasma na parede celular primária. Neste processo é determinante o aumento do volume do vacúolo.
No final da citocinese, forma-se uma delgada camada rica em pectinas – a lamela média (middle lamella) – com a função de aglutinar as células recém-diferenciadas. Nas células adultas, a lamela média posiciona-se por fora da parede primária. Tem um papel crucial na integridade dos tecidos e órgãos porque previne que as células escorreguem umas nas outras (Zamil & Geitmann, 2017).
A maioria das células parenquimatosas, incluindo as de clorênquima (parênquima com cloroplastos), mantém uma parede de natureza primária. O mesmo acontece com as células meristemáticas, os elementos crivosos e o colênquima. Os restantes tipos celulares têm parede primária enquanto jovens; concluído o período de alongamento celular, constroem uma parede secundária por dentro da parede primária. A parede celular secundária resulta da acumulação de compostos que a enrijecem, sobretudo de celulose (que pode atingir os 94% do seu peso seco), em geral combinada com lenhina, um polímero aromático complexo hidrófobo, que confere resistência estrutural, proteção contra os raios UV, melhora a condutividade hidráulica (do xilema) e participa nas defesas das plantas contra os seus inimigos. Como adiante se explica, as células da endoderme (v. «Estrutura primária da raiz») e do felema (v. «Periderme») acumulam suberina, uma outra substância hidrofóbica anti-UV, desta feita de natureza cerosa (Krizková et al., 1999).
Interrompem as paredes celulares, tanto primárias como secundárias, pequenas aberturas conhecidas por pontuações (pits). As pontuações podem ser simples, ou mostrar uma aréola (pontuações areoladas) (v. «Tecido vascular»). As pontuações de duas células vizinhas geralmente opõem-se. Nos tecidos vivos são percorridas por cordões microscópicos de protoplasma com origem no retículo endoplasmático – os plasmodesmos. A continuidade membranar e protoplasmática das células vegetais (células cenocíticas) não tem paralelo nos animais, embora não seja caso único entre os seres vivos (evoluiu de forma independente nas algas verdes e nos fungos). Os plasmodesmos permitem o transporte polar de grandes moléculas, o que facilita a nutrição de células especializadas (e.g., elementos crivosos) e o controlo fisiológico da diferenciação celular (e.g., através do transporte polar da auxina, um regulador de crescimento).
Porém, a parede celular celulósica característica das plantas verdes dificulta a excreção de subprodutos tóxicos do metabolismo. Uma das soluções evolutivas encontradas para este problema consistiu na sua acumulação num vacúolo, que, nas células mais velhas, ocupa a quase totalidade do protoplasma (~80%), e comprime o citoplasma e o núcleo para a periferia da célula. Além da acumulação de resíduos, o vacúolo foi trabalhado pela evolução para conferir rigidez estrutural às células, acumular químicos tóxicos para os inimigos das plantas, ou armazenar substâncias de reserva ou certos pigmentos hidrossolúveis (e.g., antocianinas de pétalas e folhas). O vacúolo participa ainda no processo de alongamento celular e é determinante na proteção das células contra a desidratação nas plantas vasculares.
A parede celular e o vacúolo são duas das muitas características que diferenciam a célula vegetal da célula animal, descritas ao pormenor nos livros-texto de biologia.
Outras características da célula vegetal
Além do citoplasma, núcleo e organitos celulares (inc. vacúolo) (Figura I.2.1), as células vegetais contêm, frequentemente, inclusões de vária ordem; e.g., grânulos de amido e cristais de oxalato de cálcio. Dizem-se ergásticas as substâncias de reserva ou os subprodutos do metabolismo constituintes de acumulações intracelulares ou inclusos na parede celular. As células com substâncias ergásticas caem, muitas vezes, no conceito de idioblasto (v.i.).
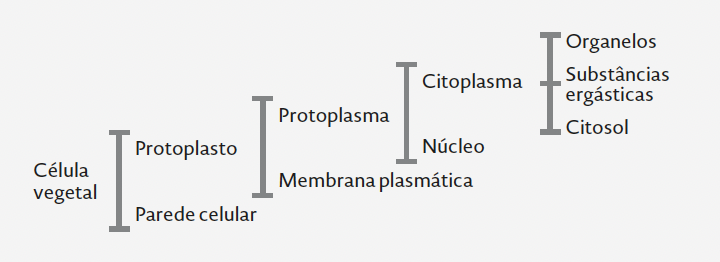 FIGURA I.2.1. Constituintes da célula vegetal. [Original.]
FIGURA I.2.1. Constituintes da célula vegetal. [Original.]
As ráfides são inclusões cristalinas alongadas em forma de agulha de oxalato de cálcio ou de carbonato de cálcio, comuns e de fácil observação microscópica, que se admite terem algum efeito na contenção da herbivoria. Por exemplo, são abundantes na Oxalis pes-caprae (Oxalidaceae), no Chlorophytum comosum (Asparagaceae, Agavoideae) e nos inhames da família das aráceas (e.g. Colocasia esculenta e Xanthosoma sagittifolium; Figura I.2.2).

FIGURA I.2.2. Ráfides. As folhas de Xanthosoma sagittifolium (Araceae) acumulam grandes quantidades de ráfides de oxalato de cálcio que se espetam facilmente na pele causando um intenso prurido. Deambular numa plantação de Xanthosoma não é uma boa ideia. As folhas e os rizomas são edíveis após cozedura. [Wikimedia Commons.]
Muitas gramíneas e cucurbitáceas, entre outras plantas, sobretudo nas células da epiderme, depositam grande quantidade de sílica, intra e/ou extracelularmente sob a forma de acumulações microscópicas conhecidas por corpos de sílica ou fitólitos (phytoliths). As células com estas propriedades são designadas por células silicificadas (silicified cells). Os fitólitos têm valor taxonómico e acumulam-se no solo – são uma valiosa fonte de informação na reconstrução paleoclimática e no estudo da domesticação das plantas (Piperno, 2006). Alguns investigadores admitem que as deposições de sílica sejam o resultado do transporte inadvertido de ácido silícico a partir do solo, uma vez que este tende a acumular-se nos tecidos vasculares e nas células epidérmicas localizadas no final da corrente respiratória (Motomura et al., 2000). Foi demonstrado em gramíneas tropicais que a formação de corpos de sílica nas folhas é maior nas plantas provenientes de pastagens sujeitas a uma herbivoria intensa e nas plantas intensamente desfoliadas. Aparentemente, a silicificação dos limbos das gramíneas é adaptativa e tem a função de deter a herbivoria (McNaughton & Tarrants, 1983). O silício tem um vasto papel de proteção das plantas contra o stresse e as radiações ionizantes (Luyckx et al., 2017).
Orientação da divisão celular
O desenvolvimento celular compreende três processos essenciais:
- Divisão e multiplicação celulares – incremento do número de células;
- Alongamento celular – aumento da dimensão das células;
- Diferenciação celular – especialização funcional das células.
As células vegetais dividem-se por mitose, outro tema abordado em detalhe nos livros de biologia geral. A meiose é um evento constante, mas raro no ciclo de vida das plantas terrestres, sempre associado com a produção de esporos (v. «Ciclos de vida das ‘plantas de esporulação livre’»). Nas plantas, a individualização das células filhas (citocinese) resultantes de mitose é de imediato sucedida pela formação de parede celular.
As células vegetais estão cingidas por uma parede celular rígida, portanto, a forma dos órgãos vegetais reflete os planos de orientação das mitoses e do crescimento celular durante o processo de desenvolvimento. As divisões celulares paralelas à superfície de um órgão – divisões periclinais (Figura I.2.3-A) – aumentam a sua espessura. As divisões anticlinais são perpendiculares à sua superfície. Nos órgãos cilíndricos (= axiais) – raiz e caule –, as divisões celulares anticlinais, por sua vez, podem ser transversas (perpendiculares ao eixo longitudinal) (Figura I.2.3-B) ou radiais (decorrem num plano longitudinal). As divisões anticlinais transversas alongam os órgãos. As divisões anticlinais radiais ocorrem, por exemplo, no câmbio vascular e na felogene (v. «Meristemas») em resposta às tensões resultantes do aumento do diâmetro de raízes ou caules causados pelo crescimento secundário.
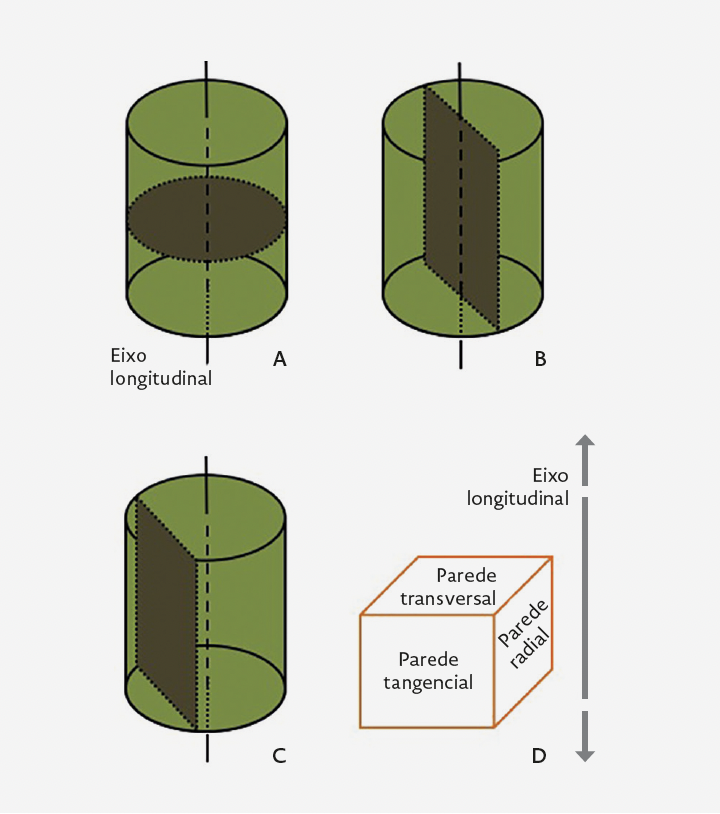
FIGURA I.2.3. Representação esquemática da orientação das divisões celulares em corte radial. A) Divisão periclinal. B) Divisão anticlinal transversa. [Original.]
[1] A fagocitose persiste na linhagem mais antiga de plantas verdes, os prasinodermófitos (Maruyama & Kim, 2013).