
2. Ciclo de vida das gimnospérmicas
Gimnospérmicas atuais
As gimnospérmicas atuais (acrogimnospérmicas) dividem-se em cinco grandes grupos (subclasses): Cycadidae, Ginkgoidae, Cupressidae, Pinidae e Gnetidae (Figura 322). Correntemente, são aceites cerca de 1045 espécies de gimnospérmicas, repartidas por 84 géneros e 12 famílias (Christenhusz et al., 2011; Roskov et al., 2019). As 'coníferas' (Pinidae + Cupressidae) incluem a maior parte dos géneros (70) e das espécies (615). Os restantes três grupos de 'não coníferas' compreendem: 317 espécies de cicadófitos (em 10 géneros repartidos por 3 famílias); um único ginkgófito vivo (Ginkgo biloba, Ginkgoaceae); e 112 gnetófitos em três famílias monogenéricas (Ephedraceae, Welwitschiaceae e Gnetaceae).
Órgãos de suporte, estruturas reprodutivas e estróbilos
Nas gimnospérmicas, os fenómenos anatómicos e fisiológicos relacionados com a reprodução sexuada decorrem em estruturas reprodutivas unissexuais. Em contraste, as angiospérmicas possuem um órgão especializado na reprodução sexual que é originalmente hermafrodita: a flor.
Nas gimnospérmicas monoicas, cada indivíduo possui, em diferentes pontos da sua copa, estruturas reprodutivas ♂ e ♀; e.g., os pinheiros (Pinus, Pinaceae) e os abetos (Abies, Pinaceae). Nas espécies dioicas, numa mesma população coexistem indivíduos exclusivamente ♂ e indivíduos ♀; e.g., o ginkgo e o teixo (Taxus baccata, Taxaceae) (Figura IV.C.2.12-A).
Nas gimnospérmicas atuais, os órgãos de suporte (de sacos polínicos ou primórdios seminais) surgem, salvo raras exceções, agregados. As estruturas reprodutivas unissexuais resultantes desta agregação arquitetónica designam-se genericamente por estróbilo (ou cone) (Figuras IV.C.2.1 e IV.C.2.1.2). Existem duas exceções: não são, geralmente, considerados estróbilos as estruturas reprodutivas ♀ das Taxaceae nem as do ginkgo (Figuras IV.C.2.12-A e IV.C.2.5).
O termo latino strobilus refere-se originalmente a estruturas reprodutivas em forma de cone, constituídas por um eixo caulinar central, no qual se inserem escamas que, na maturidade, se tornam lenhosas. Com este sentido estrito, o termo é aplicado tanto às gimnospérmicas como a alguns fetos (e.g., Equisetum). Contudo, na bibliografia botânica atual, constata-se a tendência de aplicar o termo estróbilo num sentido lato a todas as estruturas reprodutivas que aglomeram órgãos de suporte de microsporângios e/ou de primórdios seminais. Sob este prisma conceptual, também a flor das angiospérmicas pode ser definida evolutivamente como um estróbilo altamente modificado e, regra geral, bissexual.
Nas plantas com semente, o pólen forma-se e matura no interior de sacos polínicos. Nas gimnospérmicas, estes sacos polínicos dispõem-se em grupos de 2 a 12 na face abaxial (inferior) de microsporofilos foliares muito especializados —as escamas polínicas (Figuras IV.C.2.6-A,C e IV.C.2.8-A). Estes órgãos de suporte estão, por sua vez, e em número variável, sempre organizados em estróbilos ♂. Os estróbilos ♂ da esmagadora maioria das acrogimnospérmicas são anatomicamente simples (com eixos não ramificados) (Schulz et al., 2014).
Os primórdios seminais das gimnospérmicas atuais contactam diretamente o exterior — não estão encerrados num pistilo, como ocorre nas angiospérmicas. Consoante o grupo taxonómico, os primórdios seminais das gimnospérmicas apresentam-se espacialmente arrumados de quatro formas:
- Solitários, envolvidos por uma estrutura carnuda de origem foliar (arilo), como nas Taxaceae (com algumas exceções) (Figura IV.C.2.12-A);
- Aos pares (raramente solitários ou em grupos de três), na extremidade de um ramo fértil curto e bifurcado, como no Ginkgo (Figura IV.C.2.5);
- Na margem de megasporofilos pouco modificados, aglomerados em estróbilos ♀ laxos, como no género Cycas (Cycadaceae) (Figuras IV.C.2.4-A,B);
- Em estróbilos ♀ compactos/congestos, geralmente lenhosos e de estrutura anatómica muito variável; tipo mais frequente (vd. «Interpretação evolutiva do estróbilo ♀»; Figura IV.C.2.4-C,D, Figura IV.C.2.6-B,D e Figura IV.C.2.8-C,D).
As angiospérmicas têm uma camada externa extra, e exclusiva, de proteção do gametófito e do gâmeta ♀: o carpelo (um ou mais organizados em pistilos). A solução encontrada na linhagem das gimnospérmicas foi distinta, uma vez que o seu primórdio seminal (mais concretamente, o micrópilo) tem de estar fisicamente exposto ao exterior durante a polinização para captar o pólen. Assim, em grupos avançados como as araucariáceas, as 'coníferas' e as zamiáceas, o primórdio seminal está inserido (e parcialmente fundido) numa escama protetora, no âmbito de um estróbilo lenhoso. Esse mesmo estróbilo, logo após a polinização é aproveitado para encerrar ao exterior e proteger os primórdios fecundados e, mais tarde, já maduro, eventualmente auxilia a dispersão da semente.

Figura IV.C.2.1. Estrutura do estróbilo ♂ das Pinaceae. Cone ♂ de Pinus sp. (Pinaceae). N.b.: sacos polínicos preenchidos com grãos de pólen inseridos na face abaxial de escamas polínicas. [Cortesia da Cornell University Plant Anatomy Collection].
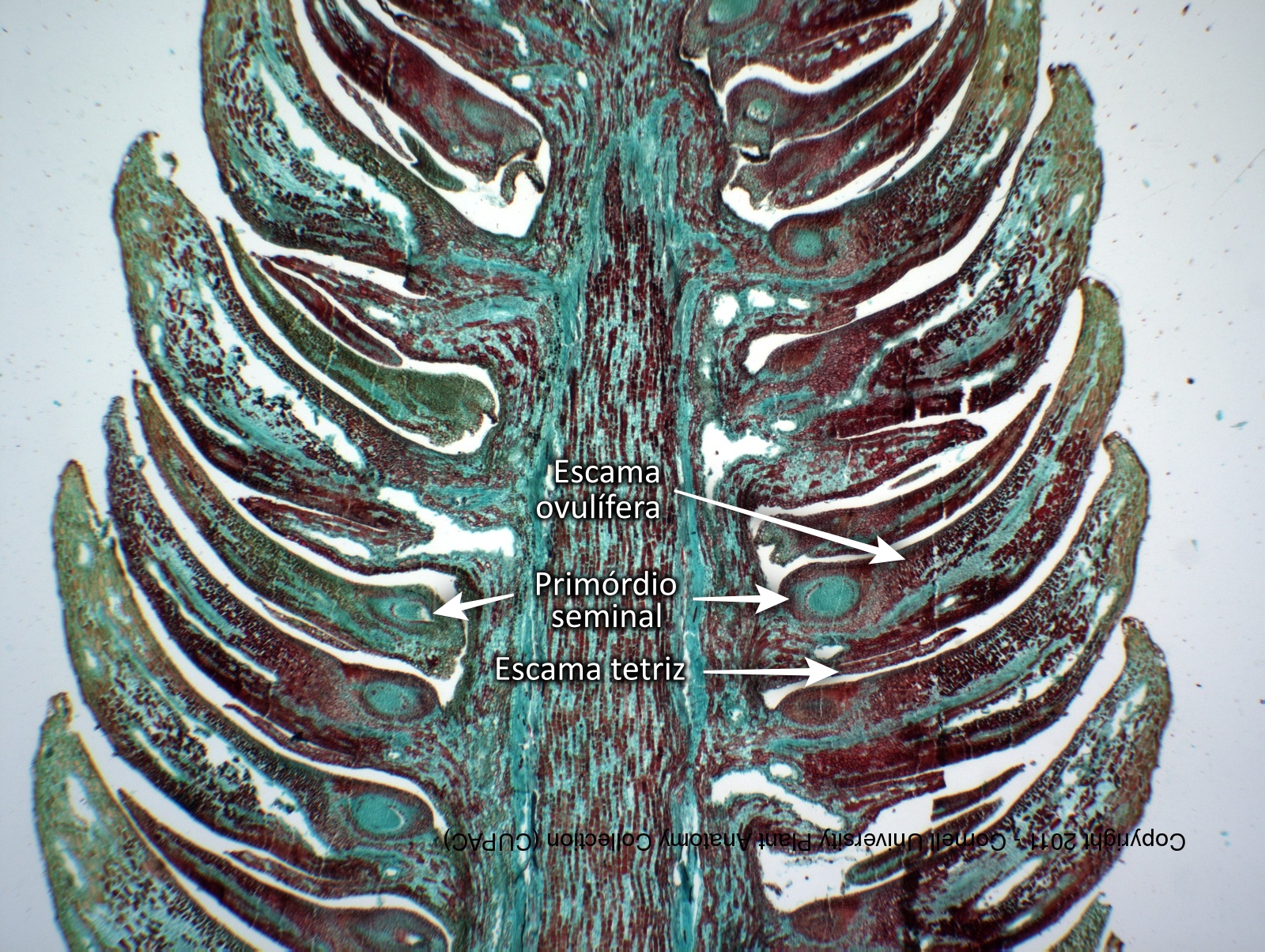
Figura IV.C.2.2. Estrutura do estróbilo ♀ das Pinaceae. Cone de Pinus wallichiana (Pinaceae). N.b.: no canto inferior esquerdo, escamas ovulíferas com um primórdio seminal apenso na base, axiladas por uma escama tetriz. [Cortesia da Cornell University Plant Anatomy Collection].
Interpretação evolutiva do estróbilo feminino
A interpretação evolutiva do estróbilo ♀ nos cinco grandes grupos de acrogimnospérmicas é complexa e controversa. Embora não se tenha ainda atingido uma interpretação de absoluto consenso, é hoje claro que os estróbilos, e as suas respetivas componentes anatómicas, nem sempre são estruturas homólogas entre os diferentes grupos.
Nas Cycadaceae (Cycadidae), a família de acrogimnospérmicas que retém mais características ancestrais (plesiomorfias), os primórdios seminais estão localizados na margem de megasporofilos que mantêm ainda uma nítida morfologia foliar. Estes megasporofilos, por sua vez, agrupam-se no topo dos caules, diferenciando um estróbilo ♀ laxo (Figura IV.C.2.4-A,B). Os primórdios seminais estão totalmente a descoberto antes da polinização, uma exposição que, nas plantas atuais, só tem equivalente no ginkgo. Nas Zamiaceae, a família mais avançada da subclasse Cycadidae, os megasporofilos estão metamorfoseados em escamas lenhosas organizadas em estróbilos muito semelhantes aos cones de algumas pináceas, com os primórdios seminais bem resguardados no seu interior (Figura IV.C.2.4-D). Os estróbilos ♀ dos cicadófitos dizem-se simples porque se circunscrevem a um único eixo com folhas modificadas (megasporofilos) apenas — não são caules ramificados.
O ginkgo segue um modelo arquitetónico completamente diferente: as estruturas reprodutivas ♀ resumem-se a um ramo fértil bifurcado (ou trifurcado), que ostenta apenas 2 ou 3 primórdios seminais na extremidade distal (número maior em algumas espécies fósseis de ginkgófitos). Supõe-se que este ramo fértil seja o resultado evolutivo da hiper-simplificação de um estróbilo ♀ ancestral.
No eixo central do estróbilo ♀ das pináceas (a única família do clado dos pinófitos) inserem-se dois tipos de componentes: a escama (ou bráctea) tetriz e a escama ovulífera (Figuras IV.C.2.2 e IV.C.2.12-B,C). Cada escama tetriz axila uma escama ovulífera; por sua vez, na face adaxial (superior) de cada escama ovulífera repousam 2 primórdios seminais (Figura IV.C.2.12-B). O botânico sueco C. R. Florin (1894-1965) propôs que a escama tetriz tem origem foliar — é uma verdadeira bráctea —, sendo a escama ovulífera um sistema caulinar modificado (não é uma bráctea) (Florin, 1951). O desenho e a dimensão relativa das escamas tetriz e ovulífera são atributos morfológicos muito valorizados na taxonomia e segregação dos géneros de pináceas (Figura IV.C.2.12-C). Portanto, o estróbilo ♀ das pináceas é, na realidade, um sistema de caules, i.e., um estróbilo composto muito simplificado, desprovido de megasporofilos (Figura IV.C.2.7-A).
Os cupressófitos são morfologicamente muito mais diversos do que os pinófitos. Comportam cinco famílias atuais: Araucariaceae, Podocarpaceae, Sciadopityaceae, Cupressaceae (que agora inclui as antigas Taxodiaceae) e Taxaceae. Na família das cupressáceas (a mais diversa do grupo), as escamas ovulíferas estão reduzidas a uma mera protuberância (invisível nas frutificações maduras) nos géneros mais basais da família (em parte das antigas taxodiáceas, e.g., Sequoia e Cryptomeria). Este resquício evolutivo caulinar desaparece por completo nos grupos mais avançados da família (as Cupressaceae s.str.). Em ambos os casos, os estróbilos ♀ das cupressáceas são constituídos por um eixo principal (por vezes muito curto) revestido por brácteas, homólogas das brácteas tetrizes dos pinófitos — natureza foliar, portanto (Figura IV.C.2.3, Figura IV.C.2.7-A). Estas brácteas (ou escamas) classificam-se como férteis ou estéreis consoante suportem, ou não, primórdios seminais. Por sua vez, as brácteas podem tornar-se lenhosas (e.g., Cupressus e Chamaecyparis) ou carnudas (e.g., Juniperus), sendo esta última modificação uma clara adaptação à dispersão das sementes por animais (zoocoria).
Nas Araucariaceae, uma família basal de cupressófitos com grande importância florestal no hemisfério Sul, os estróbilos ♀ exibem apenas brácteas férteis, na superfície das quais se inserem os primórdios seminais (Conway, 2013). Nas antigas taxodiáceas, os primórdios estão localizados na axila ou na base das brácteas: as brácteas são também férteis (Tomlinson & Takaso, 2002). Nas cupressáceas mais evoluídas (e.g., Cupressus), os primórdios seminais inserem-se em várias camadas diretamente no eixo central dos estróbilos: as brácteas são meramente estéreis e protetoras (IV.C.2.7-B). Conclui-se, então, que os estróbilos ♀ são morfologicamente compostos (ramificados) nas antigas taxodiáceas, e secundariamente simples nas Cupressaceae s.str.
Os estróbilos ♀ das pináceas e das cupressáceas evidenciam, portanto, uma ontogenia e uma morfologia de todo distintas (Figura IV.C.2.7) e, muito provavelmente, nem sequer partilham uma ancestralidade comum direta. Alguns autores estendem abusivamente o conceito de megasporofilo às brácteas férteis das cupressáceas e de outras famílias de cupressófitos, enquanto outros consideram que não há quaisquer evidências destas estruturas foliares nos estróbilos das gimnospérmicas atuais. O debate evolutivo continua em aberto. Para evitar inconsistências, neste manual e no âmbito das gimnospérmicas, apenas se aplica o termo megasporofilo aos cicadófitos.
A tendência evolutiva para a simplificação do estróbilo ♀ ocorreu de forma convergente e independente em várias linhagens de 'coníferas'. O estróbilo dos zimbros (Juniperus) possui apenas de 1 a poucos primórdios. As Podocarpaceae, uma família com alguma expressão na África tropical e Australásia, retêm apenas um órgão de suporte de primórdios seminais (homólogo da escama ovulífera das pináceas) denominado epimácio, por baixo do qual se insere um número variável de brácteas estéreis, algumas das quais se tornam carnudas e coloridas na maturação (Conway, 2013; Figura IV.C.2.11). As Taxaceae, uma família muito avançada de cupressófitos, representam o caso mais extremo de simplificação do estróbilo ♀: nos teixos (Taxus), diferenciam-se apenas primórdios seminais solitários, encapsulados basalmente por um arilo carnudo na extremidade de ramos curtos. Embora morfologicamente estas estruturas de Ginkgo e Taxus não se assemelhem a estróbilos, os estudos filogenéticos mostram que os ancestrais de ambas as famílias possuíam estróbilos complexos. Por essa razão, a taxonomia moderna considera que todas as estruturas reprodutivas das gimnospérmicas, quer ♀ quer ♂, são, na sua essência evolutiva, estróbilos.
O grupo mais derivado das acrogimnospérmicas, os gnetófitos, reparte-se por três famílias de plantas dioicas (com raras exceções) de morfologia extremamente díspar. Os gnetófitos possuem estróbilos compostos unissexuais, cuja complexidade morfológica não será aqui discutida ao pormenor (Figura IV.C.2.9). Pontualmente, algumas espécies do género Gnetum (Gnetaceae) produzem estróbilos bissexuais (embora funcionalmente ♂) constituídos por anéis com vários verticilos de microsporofilos, distalmente rematados por verticilos de primórdios seminais estéreis. Nestes casos, a gota de polinização segregada pelos primórdios estéreis evoluiu para uma recompensa açucarada destinada a atrair insetos polinizadores (Endress, 1996). A tendência para aproximar fisicamente os esporângios ♀ e ♂, embora sem a criação de verdadeiros estróbilos bissexuais funcionais, repete-se na Welwitschia (Endress, 1996). Os estróbilos ♀ destas plantas, porém, têm sempre e apenas verticilos de primórdios seminais. Nos gnetófitos, tanto as estruturas de suporte dos sacos polínicos como os próprios primórdios seminais têm na sua base brácteas fundidas que formam uma estrutura análoga a um perianto floral (Kubitzki, 1990).
As múltiplas convergências evolutivas morfológicas e ecológicas dos gnetófitos com as angiospérmicas (recorde-se que as flores são um estróbilo bissexual) foram, durante quase um século, uma fonte inesgotável de equívocos evolutivos. Entretanto, a filogenómica moderna veio demonstrar cabalmente que os gnetófitos, ao contrário do que se supunha, não são os parentes evolutivos mais próximos das angiospérmicas (Ran et al., 2018).

Figura IV.C.2.3. Natureza foliar das brácteas do estróbilo ♀ das Cupressaceae: gálbulos teratogénicos de Cryptomeria japonica (Cupressaceae, Cupressidae). Os meristemas que deram origem a estes gálbulos reverteram para uma condição vegetativa após diferenciar as brácteas do estróbilo ♀. Provavelmente, mantiveram-se funcionais até que a morte programada das células dos estróbilos maduros interrompeu as suas conexões vasculares com o ramo onde se inserem. [Ilha de S. Miguel, Açores; fotografia do autor].
A) B)
B)
C) D)
D)
Figura IV.C.2.4. Estruturas reprodutivas de Cycadidae. A) Cycas revoluta (Cycadaceae), estróbilo ♀ imaturo. B) Mesma espécie, megasporofilo com primórdios seminais na base. C e D) Encephalartos munchii (Zamiaceae), estróbilos ♂ e ♀, respetivamente; espécie endémica de Moçambique polinizada por coleópteros curculionídeos. [A e B) fotografias do autor; C e D) cortesia de Pedro Capela].

Figura IV.C.2.5. Estruturas reprodutivas ♀ de Ginkgo biloba (Ginkgoaceae, Ginkgoidae). N.b.: órgãos de suporte ♀(neste caso ramos férteis) com dois primórdios seminais, inseridos na base de folhas em forma de leque. [Fotografia do autor].
A) B)
B)
C) D)
D)
Figura IV.C.2.6. Estruturas reprodutivas de Pinaceae (Pinidae). A) Estróbilos ♂ de Pinus pinaster aglomerados em grande número ao longo de um ramo do ano. B) Estróbilo ♀ da mesma espécie pouco depois da polinização, com os primórdios seminais isolados do exterior em consequência do crescimento intercalar das escamas ovulíferas. C) Estróbilos ♂ de Cedrus atlantica; n.b.: separação das escamas polínicas e deiscência do pólen de cima para baixo. D) Na mesma espécie, estróbilos ♀; n.b.: grãos de pólen aderidos às escamas ovulíferas. [Fotografias do autor].
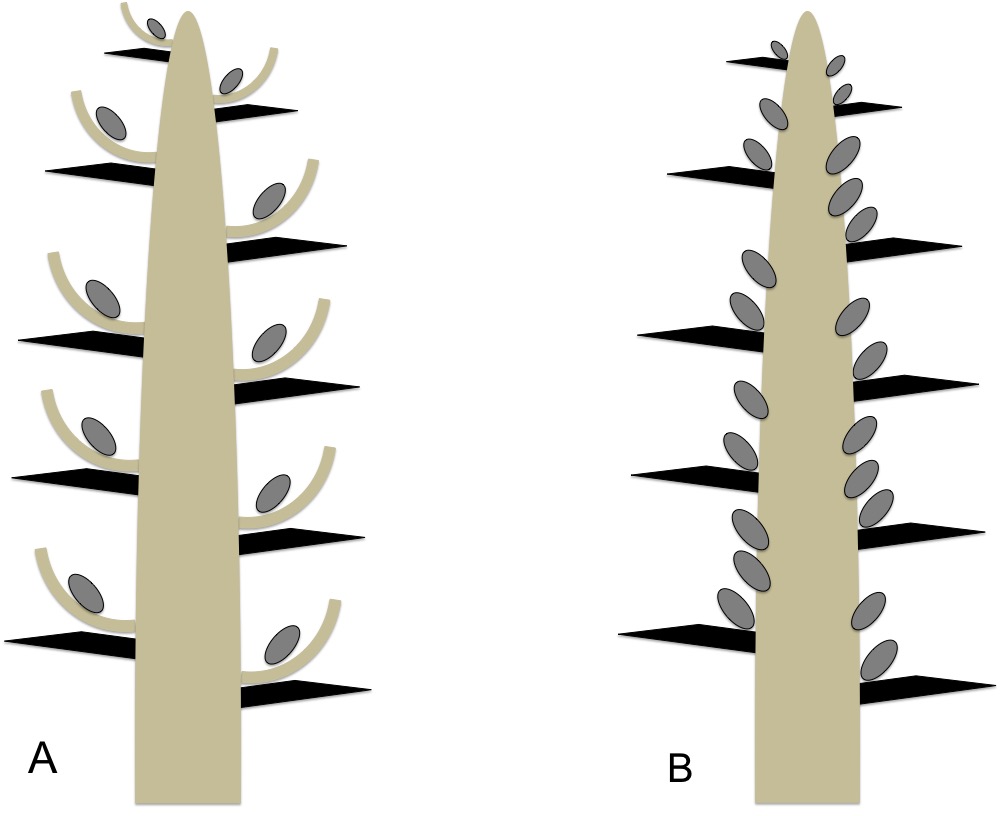
Figura IV.C.2.7. Representação diagramática do estróbilo ♀ composto e secundariamente simples nas coníferas (em corte longitudinal). A) Estróbilo ♀ composto de Pinaceae. B) Estróbilo ♀ secundariamente simples de Cupressaceae s. str.. Estruturas caulinares a castanho; estruturas foliares (brácteas) a negro; primórdios seminais a cinzento. [Original].
A) B)
B)
C) D)
D)
Figura IV.C.2.8. Estruturas reprodutivas de Cupressaceae (Cupressidae). A) Estróbilos ♂ de Chamaecyparis lawsoniana com sacos polínicos por abrir inseridos na face dorsal de microsporofilos. B) Estróbilo ♀ da mesma espécie; n.b.: primórdios seminais livres em várias camadas na axila das brácteas, com uma gota de polinização exsudada pelo micrópilo; as brácteas depois de maduras dão origem às escamas dos gálbulos (Figura IV.C.2.12-D). C) Estróbilos ♂ de Juniperus oxycedrus; n.b.: posição axilar dos estróbilos, enquanto em Chamaecyparis são terminais. D) Estróbilos ♀ da mesma espécie; n.b.: bractéolas a revestir o caule que suporta o estróbilo; as brácteas são carnudas na maturação (Figura IV.C.2.12-E). [Fotografias do autor].
Figura IV.C.2.9. Estruturas reprodutivas dos gnetófitos. A) Gnetum africanum (Gnetaceae): 1 e 4 – hábito; 2 – pormenor do estróbilo ♂; 3 – microsporofilo com várias brácteas na base; 5 – estróbilo ♀; 6 – semente. B) Gnetum gnemon: estróbilo ♂ com verticilos de microsporofilos encimados por verticilos de primórdios seminais estéreis que, apesar de tudo, produzem uma gota de polinização (visível na figura). C) Ephedra fragilis subsp. fragilis (Ephedraceae): pormenor de estróbilo ♀ com primórdios seminais revestidos com brácteas; a hipótese de que as peças bracteolares de uma gnetácea estão na origem do perianto floral é tentadora, mas falsa (volume II). D) Na mesma espécie, estróbilos maduros constituídos, neste caso, por uma semente madura rodeada por brácteas carnudas e vermelhas; n.b.: caules articulados. [A)http://www.prota.org; B) cortesia de Kevin C. Nixon e http://www.plantsystematics.org; C e D) cortesia de Miguel Porto e de Flora-on].
Biologia da reprodução
O ciclo de vida das plantas do género Pinus é, muitas vezes e de forma muito redutora, assumido como a referência estrutural e evolutiva para todas as gimnospérmicas (Figura IV.C.2.13). É importante ter presente que, embora formem um grupo monofilético, as gimnospérmicas atuais constituem um conjunto de plantas com semente extremamente heterogéneo e antigo (muito anterior à emergência das plantas com flor), com clados que divergiram entre si há centenas de milhões de anos. Nos parágrafos que se seguem, resumem-se algumas das diferenças da biologia da reprodução, e implicitamente, do ciclo de vida, que separam os cinco grandes grupos de gimnospérmicas atuais: Cycadidae, Ginkgoidae, Cupressidae, Pinidae e Gnetidae. Mas antes de prosseguir, recomenda-se uma leitura atenta do ciclo genérico da Figura IV.C.2.13.
Micro e megasporogénese. Primórdio seminal
Os grãos de pólen (n) diferenciam-se por meiose, e em enorme número, a partir de microsporócitos (2n) no interior de sacos polínicos. Nas angiospérmicas, o micrósporo sofre uma primeira mitose (seguida frequentemente de uma segunda mitose restrita à célula generativa), pelo que o pólen é libertado com 2 ou 3 células e, no momento da fecundação, o gametófito ♂ tem invariavelmente 3 células. As contas são mais complexas nas gimnospérmicas, porque o seu gametófito ♂ (pelo menos no momento da fecundação) é bastante menos reduzido do que o das angiospérmicas. O pólen é libertado com 1 a 5 células, com a notável exceção das podocarpáceas, cujo pólen pode ultrapassar as 40 células no momento da dispersão. O grão de pólen das Taxaceae, por outro lado, é libertado contendo apenas uma célula; para todos os efeitos, é um micrósporo puro, um caso único entre as plantas com semente (Anderson & Owens, 2000).
No momento da libertação dos gâmetas (i.e., já às portas dos arquegónios), o microgametófito maduro tem, por exemplo, 3 células nas Cupressaceae, 4 nas Taxaceae, 5 nas Cycadidae, 6 no Ginkgo e nas Pinaceae, e dezenas nas Podocarpaceae (Fernando et al., 2009). Ao invés do que sucede nas angiospérmicas, os especialistas conseguem ainda reconhecer no microgametófito das gimnospérmicas (exceto nas cupressáceas) resíduos evolutivos do protalo (1 ou 2 células protaliais estéreis) e do anterídio (1 célula anteridial). Contudo, à semelhança das angiospérmicas, ocorre sempre a diferenciação de uma célula do tubo e de uma célula generativa (que dará origem, após a polinização, a 2 gâmetas ♂, com raras exceções) (Fernando et al., 2009).
Como será detalhado no volume II, nas plantas com semente o saco embrionário (megagametófito ou gametófito ♀) resulta da germinação de um megásporo confinado pelas próprias paredes do esporo (endomegasporia). Por seu turno, o gametófito ♀ germinado fica retido de forma permanente (endoprotalia) no interior do megasporângio (nucelo). Adicionalmente, megasporângio é externamente envolvido por um ou dois tegumentos de origem caulinar (um nas gimnospérmicas, um ou dois nas angiospérmicas), constituindo um primórdio seminal. Esta acumulação de camadas de proteção sobre os primórdios seminais é uma forte tendência evolutiva partilhada pelas plantas e pelos animais (Grant, 1950).
Ao contrário do que se verifica nas angiospérmicas, na esmagadora maioria das gimnospérmicas a diferenciação funcional dos megasporócitos (2n) depende do estímulo químico da polinização: é, portanto, posterior a esta. O megasporócito gera, por meiose, 4 megásporos (n), um dos quais (geralmente, o calazal, i.e., o mais afastado do micrópilo) sobrevive, germina e dá origem ao gametófito ♀. O gametófito ♀ maduro das gimnospérmicas contém milhares de células e diferencia múltiplos arquegónios: 2 a 5 nas 'coníferas', geralmente em muito maior número nas Ginkgoidae e Cycadidae, e 1 a 3 em Ephedra (o mais basal dos três géneros de Gnetidae) (Fernando et al., 2009). Curiosamente, as células do gametófito ♀ das Ginkgoidae são clorofiladas (verdes). Cada arquegónio tem à sua custódia uma única oosfera. O desenvolvimento de arquegónios típicos, contudo, não é claro em dois géneros muito derivados de Gnetidae — Welwitschia (Figura 7) e Gnetum —, tal como, aliás, acontece nas angiospérmicas (Carmichael & Friedman, 1995).
Nas Ginkgoidae e nas Cycadidae, o primórdio seminal tem a mesma dimensão da semente madura, que é atingida logo depois da polinização, mas antes da fecundação ocorrer. Este padrão de crescimento constitui um investimento reprodutivo de elevadíssimo risco energético, pois, se o primórdio não for fecundado e abortar, a planta deita a perder um imenso investimento em biomassa e reservas. As 'coníferas' (Cupressidae + Pinidae) são mais eficientes porque, embora também acumulem reservas no gametófito ♀ antes da fecundação, a maior parte desta acumulação é, geralmente, desencadeada pela polinização. As 'coníferas' conseguem ainda abortar precocemente primórdios não polinizados, realocando atempadamente parte da sua energia. As angiospérmicas, contudo, otimizaram este processo ao extremo da eficiência, uma vez que só começam a acumular reservas no endosperma (através da dupla fecundação) se a singamia for bem-sucedida (Baroux et al., 2002).
Polinização
Embora, por causa dos Pinus, a monoicia seja a condição mais comum na natureza em termos de biomassa, a maioria das espécies de gimnospérmicas é, na verdade, dioica. A dioicia é um mecanismo radical para forçar a polinização cruzada. Como não possuem os sistemas genéticos de autoincompatibilidade pré-zigótica das angiospérmicas, as gimnospérmicas monoicas reduzem o risco de autopolinização posicionando as estruturas reprodutivas ♀ nos ramos mais altos da canópia, por cima das estruturas ♂ (Niu et al., 2016).
Os grãos de pólen são transportados massivamente pelo vento (polinização anemófila), sendo diretamente capturados pelos primórdios seminais, os quais, para o efeito, secretam para o exterior do micrópilo uma gota de polinização, frequentemente visível a olho nu (Figura IV.C.2.8-B). Nas 'coníferas', a conjugação aerodinâmica da forma dos ramos, folhas e estróbilos gera pequenos vórtices de ar que conduzem os grãos de pólen diretamente em direção ao micrópilo dos primórdios seminais (Niklas, 1984). As Cycadidae e as Gnetidae são, contudo, secundariamente polinizadas por insetos (e.g., coleópteros nas cicas). Em algumas espécies destes dois grupos, a própria gota de polinização funciona como recompensa alimentar açucarada.
Os açúcares dissolvidos na gota de polinização reduzem as perdas de água por evaporação e, ao aumentarem drasticamente a viscosidade do líquido, diminuem a probabilidade da gota se soltar do micrópilo pela ação direta do vento e da agitação mecânica dos ramos (Lunau, 2002). Ainda assim, a folhagem de géneros como Chamaecyparis e Cupressus fica frequentemente pegajosa devido à acumulação de gotas que se destacam dos primórdios seminais no pico do período de polinização. Os grãos de pólen afundam-se na gota de polinização (ou são arrastados ativamente quando esta é fisiologicamente reabsorvida pela planta) e entram no primórdio. Em seguida, a porção da gota que fica exposta ao exterior seca e sela hermeticamente o micrópilo. O microgametófito fica assim protegido no interior da câmara polínica (Chamberlain, 1935), num ambiente mantido mais ou menos assético pela ação de proteínas antimicrobianas previamente dissolvidas na gota de polinização (Poulis et al., 2005).
Fase progâmica e fecundação
No interior da câmara polínica, o pólen germina na superfície do nucelo, rompe a esporoderme (exina) e diferencia um tubo polínico. O tubo penetra o nucelo de forma intercelular (digerindo a lamela média), ao mesmo tempo que o microgametófito vive uma sequência final de mitoses que desemboca na formação dos gâmetas ♂. O tubo polínico tem de vencer uma barreira mecânica de 0,1 a 3 mm de espessura de nucelo, um tecido que já não diferencia uma barreira nas angiospérmicas (Williams, 2008). O tubo polínico das gimnospérmicas mantém uma continuidade citoplásmica total entre a sua extremidade de crescimento e o grão de pólen original; por essa razão, é muito mais curto do que o tubo das angiospérmicas (Fernando et al., 2009). Em contraste, no tubo polínico das angiospérmicas diferenciam-se, a cada passo do seu avanço, tampões isolantes de calose que mantêm as células espermáticas e o citoplasma ativo sequestrados apenas na extremidade distal do tubo, permitindo que este atinja comprimentos assinaláveis (e.g., descendo ao longo de todo o estilete do milho) (Lord, 2000).
Nas Ginkgoidae e nas Cycadidae, o tubo polínico (que tem uma função meramente haustorial/alimentar) rompe-se na proximidade da câmara arquegonial: cada tubo liberta dois gâmetas flagelados (anterozoides) que nadam ativamente no fluido rumo às oosferas (zooidogamia ou sifonogamia imperfeita). A observação de gâmetas ♂ flagelados nestas plantas de semente estabeleceu uma conexão evolutiva entre os 'pteridófitos' e as gimnospérmicas, sendo considerada uma das mais importantes descobertas científicas botânicas do século XIX. Nas Cupressidae, Pinidae e Gnetidae — i.e., no grosso das gimnospérmicas atuais —, o tubo polínico conduz e coloca as células espermáticas imóveis em contacto direto com as oosferas (sifonogamia verdadeira).
No Ginkgo biloba, o intervalo que medeia entre a polinização e a fecundação é de 4 a 5 meses, caindo em seguida os primórdios recém-fecundados ao solo (Del Tredici, 2007). Este intervalo ronda os 6 a 12 meses nas Cycadidae, e pode estender-se de 3 meses a uns impressionantes 24 meses nalgumas 'coníferas' (Singh, 1978 cit. em Williams, 2009). Nas Gnetidae, a dinâmica é muito mais rápida: decorre apenas 1 semana em Gnetum e 10 a 36 horas em Ephedra (Fernando et al., 2009). Cada oosfera é fecundada por um único núcleo espermático. Em dois géneros de Gnetidae (Gnetum e Ephedra) foi descoberto um esboço evolutivo de dupla fecundação; contudo, o tecido nutritivo de reserva permanece invariavelmente haploide, tal como nos restantes grupos de gimnospérmicas, não se formando um endosperma verdadeiro (Carmichael & Friedman, 1995; Friedman, 2014).
Na maior parte das espécies atuais — excetuando o género Cycas, o Ginkgo e a maioria das Taxaceae —, logo após a polinização, os primórdios fecundados, como se referiu, são rapidamente envolvidos pelo crescimento de estruturas adjacentes provenientes do estróbilo ♀(escamas), as quais os isolam do exterior e os protegem de agentes bióticos e abióticos durante o longo desenvolvimento do embrião (Figura IV.C.2.6-B).
Poliembrionia
A existência comum de múltiplos arquegónios num único megagametófito gera frequentemente mais do que um embrião viável (poliembrionia simples; simple polyembryony) e, implicitamente, vários embriões em cada primórdio seminal. Este fenómeno é quase universal nas gimnospérmicas. Sobretudo em alguns géneros de Pinaceae, ocorre também, e com grande frequência, um outro tipo de embrionia, a poliembrionia por clivagem (cleavage polyembryony), que se verifica quando um único embrião jovem se cinde vegetativamente em vários outros embriões idênticos. Em Pinus, é comum o início da formação de até 8 embriões. Em ambos os sistemas, porém, a competição interna dita que geralmente apenas um embrião sobrevive até à fase final — as sementes maduras de gimnospérmicas raramente contêm mais do que um embrião viável.
Não se conhecem nas gimnospérmicas sistemas genéticos de autoincompatibilidade pré-zigótica (antes da fecundação) nem mecanismos de seleção de gâmetas (competição de tubos polínicos) similares aos que ocorrem no estilete das angiospérmicas (vd. «Sistemas de autoincompatibilidade» e «Competição do pólen. Seleção de gâmetas»). Foi proposto que a poliembrionia simples e por clivagem funciona, nestas plantas, como um mecanismo alternativo de seleção (pós-zigótico) da melhor descendência (Zavada & Taylor, 1986). Este sistema de triagem competitiva atua internamente enquanto os embriões são ainda muito pequenos, mas, ainda assim, é substancialmente menos eficiente do que os sistemas pré-zigóticos das angiospérmicas e acarreta custos energéticos incomparavelmente superiores para a planta materna.
Semente
As gimnospérmicas não têm cotilédones com função de reserva, não produzem endosperma e o nucelo (i.e., o megasporângio diploide materno, 2n) tem um papel transitório na nutrição dos tubos polínicos e na diferenciação do gametófito ♀ (na semente madura da esmagadora maioria das gimnospérmicas, o nucelo resume-se a um vestígio ensanduichado entre o tegumento e o corpo do gametófito ♀). As reservas nutritivas da semente são exclusivamente constituídas por tecido gametofítico (haploide) de origem materna, i.e., pelo próprio saco embrionário (gametófito ♀ ou megagametófito). Portanto, durante a germinação, o embrião das gimnospérmicas consome o gametófito ♀. Estão em causa um tecido de reserva e um processo evolutivo sem paralelo nas angiospérmicas. Os modernos manuais de botânica evitam a designação obsoleta de endosperma primário para o gametófito ♀, porque esta cria uma falsa ideia de homologia morfológica e evolutiva (v. Raven et al., 2014; Simpson, 2019).
O intervalo total entre a polinização e a maturação final das sementes ronda 1 ano na grande maioria das gimnospérmicas. É um pouco mais curto no Ginkgo biloba (Del Tredici, 2007). Em algumas espécies de Cycas (Cycadidae), o processo estende-se por impressionantes 2,5 anos (Pant, 2002). As sementes dos pinheiros (Pinus) amadurecem tipicamente em 2 anos (e.g., pinheiro-bravo, Pinus pinaster) ou até 3 anos (e.g., pinheiro-manso, Pinus pinea). É igualmente frequente as sementes caírem ao solo de forma ainda imatura (e.g., Ginkgo, cicas e Gnetum), terminando o desenvolvimento embrionário já na folhada. Como se referiu, as angiospérmicas reproduzem-se, por oposição, com uma enorme rapidez. No Ginkgo e nas Cycadidae, ao contrário das 'coníferas', as sementes maduras não suportam a desidratação — são sementes recalcitrantes. As sementes das gimnospérmicas apresentam geralmente dois cotilédones embrionários (embora sejam numerosos verticilados no género Pinus; Figura IV.C.2.10) e são extremamente ricas em lípidos e proteínas.
As sementes do ginkgo e dos teixos diferenciam-se e amadurecem sem qualquer proteção estrobilar lenhosa. As sementes de ginkgo desenvolvem uma sarcotesta carnuda e espessa, com um odor desagradável a ácido butírico na maturação (assemelhando-se a manteiga rançosa) — razão pela qual o planeamento urbano evita a plantação de árvores ♀ em passeios e jardins. Os ancestrais dos ginkgos foram certamente dispersos endozoocoricamente por dinossauros herbívoros no Mesozoico e pela megafauna mamífera no Plistocénico (Del Tredici, 1989). Hoje em dia, o Ginkgo biloba (a única espécie atual sobrevivente de toda a sua subclasse) é um fóssil vivo que depende do Homem para se propagar e dispersar (antropocoria). Nos teixos, as sementes nuas surgem basalmente envolvidas por uma estrutura carnuda e vermelha (arilo), muito doce e comestível para as aves (embora a semente em si, e toda a folhagem da árvore, sejam mortalmente tóxicas para a maioria dos mamíferos). O arilo atua assim como a recompensa perfeita para a dispersão endozoocórica (Figura IV.C.2.12-A). Anatomicamente, esta estrutura tem origem foliar (resulta da fusão de duas folhas modificadas), não cabendo, num sentido botânico estrito, no conceito clássico de arilo (Dörken et al., 2019).

Figura IV.C.2.10. Cotilédones nas gimnospérmicas. Plântula de pinheiro-bravo com 11 cotilédones. [Cortesia de Nuno Gomes].
Frutificações
Designa-se por frutificação (Figura IV.C.2.12) o estróbilo ♀ maduro, mono ou plurispérmico, das gimnospérmicas. O fruto, num sentido botânico estrito, resulta exclusivamente do desenvolvimento das paredes de um ovário após a fecundação (tem origem carpelar): o fruto é, portanto, uma sinapomorfia (característica exclusiva) das angiospérmicas. As frutificações lenhosas das gimnospérmicas são, assim, estruturas análogas (e não homólogas) aos frutos secos das plantas com flor.
Nas 'coníferas' reconhecem-se dois tipos fundamentais de frutificações:
- Gálbulos (ougálbulas): estróbilos de forma tendencialmente globosa, compostos por brácteas normalmente peltadas (em forma de guarda-chuva ou escudo) que se inserem mais ou menos no mesmo ponto do eixo central; e.g., frutificações lenhosas dos ciprestes (Cupressus, Cupressaceae). Apresentam um subtipo ecológico particular: os gálbulos baciformes — gálbulos de brácteas intimamente fundidas e carnudas, morfologicamente muito semelhantes a uma baga ou drupa (funcionando como tal para atrair aves), característicos dos zimbros (Juniperus, Cupressaceae) e de membros das Podocarpaceae;
- Pinhas: estróbilos alongados ou cónicos, compostos por escamas ovulíferas lenhosas imbricadas (e.g., pinhas de pinheiro [Pinus, Pinaceae]) ou brácteas espessas (e.g., cones das araucárias [Araucaria, Araucariaceae]) que se inserem ao longo de um eixo central desenvolvido.

Figura IV.C.2.11. Cone ♀ de Podocarpaceae (Pinidae). Cone maduro de Podocarpus angustifolius, uma árvore endémica de Cuba. O cone das podocarpáceas tem geralmente apenas um primórdio; cada primórdio está envolvido por uma escama ovulífera carnuda, o epimácio. Algumas brácteas situadas na base do cone são carnudas na maturação, muitas vezes vermelhas, para atraírem dispersores. [Fotografia do autor].
Figura IV.C.2.12. Frutificações das 'coníferas'. A) Semente solitária de teixo (Taxus baccata, Taxaceae) envolvida por um arilo vermelho; n.b.: bractéolas a revestir o caule que suporta a semente. B) Estrutura de uma pinha de pinheiro-manso (Pinus pinea, Pinaceae). C) Pinha de pseudotsuga (Pseudotsuga menziesii, Pinaceae); n.b.: brácteas tetrizes exsertas na axila das quais se inserem escamas ovulíferas arredondadas, cada uma com duas sementes apensas (não visíveis na fotografia). D)Gálbulos de cipreste-do-óregon (Chamaecyparis lawsoniana, Cupressaceae); n.b.: sementes aladas na axila de brácteas peltadas (em forma de guarda-chuva). E) Gálbulos baciformes de sabina-das-praias (Juniperus turbinata, Cupressaceae). [Fotografias do autor].
Dispersão e dormência
A dispersão das 'coníferas' faz-se, sobretudo, sob a forma de semente libertada a partir de estróbilos deiscentes. Nos abetos (Abies) e nos cedros (Cedrus), os estróbilos lenhosos maduros desintegram-se completamente na própria árvore, libertando-se em simultâneo do eixo central (e caindo ao solo) as escamas ovulíferas e as sementes. Nas restantes espécies de pináceas, as escamas ou brácteas do estróbilo abrem higroscopicamente na maturação (afastando-se do eixo) para que a semente possa ser arrastada e dispersa pela força do vento (anemocoria). Algumas frutificações de gimnospérmicas (particularmente no género Pinus) são marcadamente serotinas, i.e., mantêm-se seladas com resina durante anos e apenas abrem as escamas para dispersar as sementes em resposta a um estímulo térmico extremo, concretamente pela ação da passagem do fogo de um incêndio florestal (como acontece com os cones fechados do pinheiro-bravo) (Tapias et al.,2001).
Para auxiliar a sua aerodinâmica, géneros como Araucaria, Agathis, Pinus, Cedrus, Tsuga, Abies, Picea e tantos outros clados de coníferas, desenvolvem sementes aladas, cuja asa membranosa não é mais do que o resultado da expansão lateral das camadas superficiais do tegumento durante a maturação. Uma notável adaptação à anemocoria por semente alada ocorre também na Welwitschia mirabilis [Gnetidae], que usa o vento do deserto da Namíbia para dispersar as suas sementes.
Por outro lado, nas 'coníferas' que desenvolveram frutificações de textura carnuda, não se dispersa a semente nua; dispersa-se a frutificação inteira através da ingestão e transporte por animais frugívoros (zoocoria). Exemplos clássicos são os já referidos zimbros (Juniperus, Cupressaceae), os Cephalotaxus (Taxaceae) e as espécies de Podocarpaceae. Estruturas carnudas de origem foliar (brácteas modificadas) de ajuda à dispersão zoocórica ocorrem também nos gnetófitos dos géneros Ephedra e Gnetum. Da mesma forma ecológica, as sementes primitivas dos cicadófitos, por estarem providas de uma espessa e nutritiva sarcotesta, são ativamente procuradas e dispersas por mamíferos terrestres ou morcegos frugívoros, havendo também espécies insulares cujas sementes esponjosas flutuam eficazmente na água do mar (hidrocoria) (Dehgan & Yuen, 1983).
À semelhança das angiospérmicas, as sementes de muitas gimnospérmicas não germinam imediatamente após a dispersão, recorrendo a diversos mecanismos de dormência para sincronizar a germinação e a emergência com condições ambientais propícias (a maior parte das espécies temperadas e mediterrânicas exibe uma dormência fisiológica quebrada pelo frio).
Ciclo de vida das gimnospérmicas
A Figura IV.C.2.13 resume o ciclo de vida dos briófitos, tomando como modelo o pinheiro-bravo (Pinus pinaster, Pinaceae).
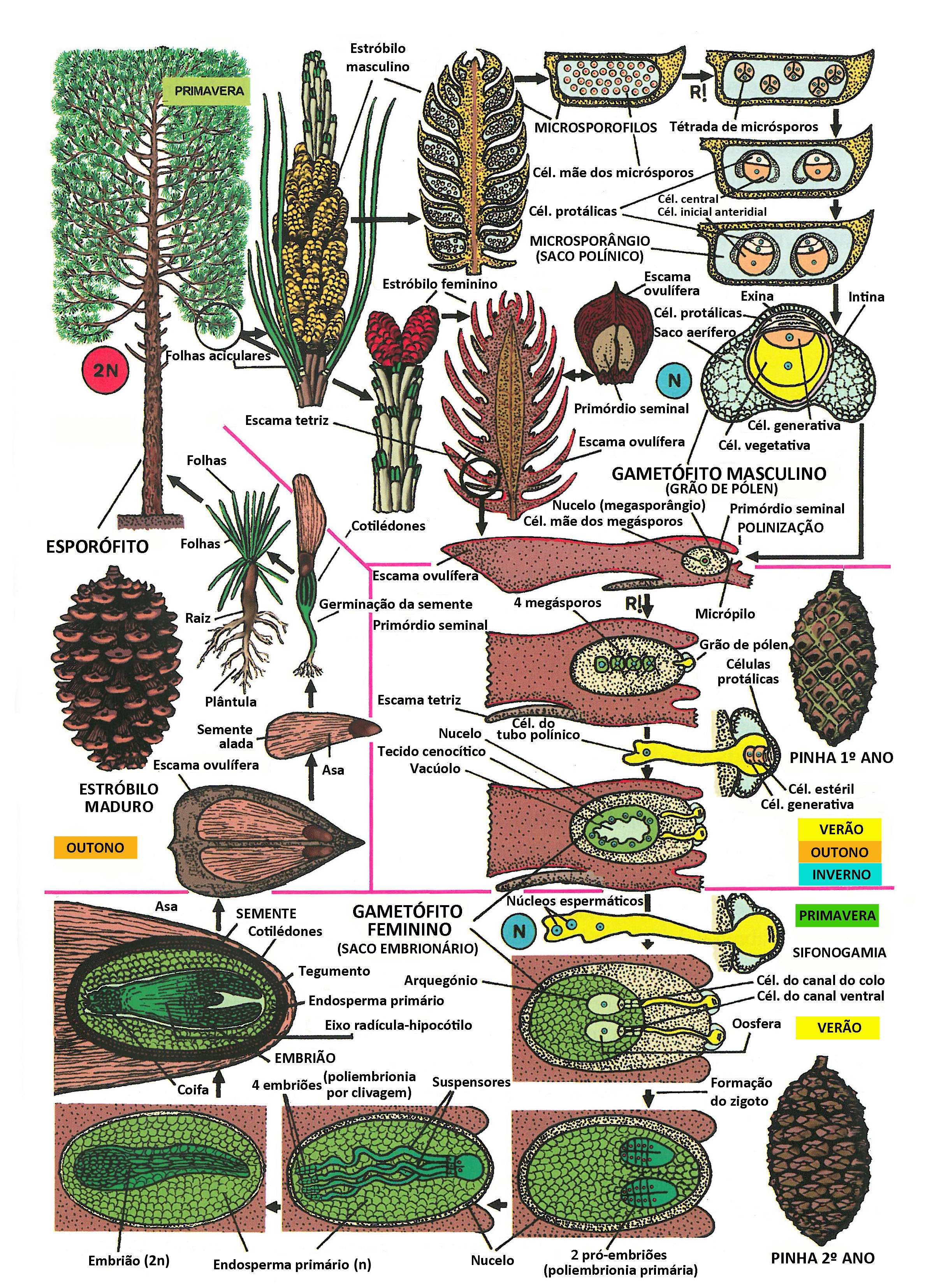
Figura IV.C.2.13. Ciclo de vida das Gimnospérmicas (Pinus pinaster, Pinaceae, Pinidae). Nomenclatura das células do gametófito ♂ de acordo com Fernando et al. (2009). Consoante se refere no texto, endosperma primário é uma designação obsoleta do megametófito. Legenda: R! – meiose. [Tradução e adaptação autorizada de um original de Tomás Díaz González, Univ. Oviedo, Espanha].