
1. Ciclo de vida dos espermatófitos atuais: estudo comparado
As gimnospérmicas e as angiospérmicas são espermatófitos, i.e., plantas com semente. As primeiras gimnospérmicas evoluíram no final do Devónico (ca. 385 Ma) (Gerrienne et al., 2004). Os cinco grandes grupos de gimnospérmicas atuais (acrogimnospérmicas) têm provavelmente um ancestral comum que datará do Carbonífero (ca. 300 M.a.) (Figura 322). As plantas com flor são mais tardias; evoluíram de uma gimnospérmica desconhecida pertencente a um grupo anterior às acrogimnospérmicas, no Triássico ou no Jurássico (no intervalo 256-149 M.a.). O ancestral de todas as angiospérmicas atuais terá vivido no Jurássico Superior (Barba-Montoya et al., 2018). No ponto «Anatomia das madeiras (breve referência)», vimos que a anatomia do lenho das gimnospérmicas e das angiospérmicas é distinta. Essas diferenças são ainda mais pronunciadas ao nível reprodutivo e na estrutura do ciclo de vida. As acrogimnospérmicas e as angiospérmicas divergem evolutivamente há muito tempo.
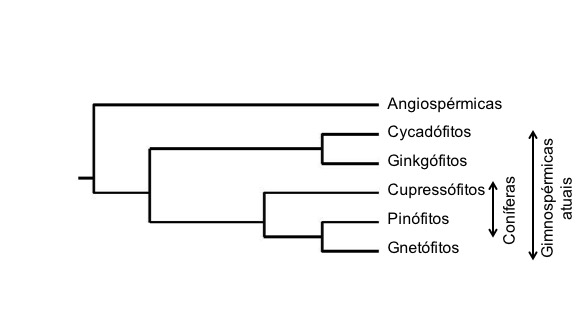
Figura 322. Relações evolutivas entre os seis grandes grupos de plantas com semente. A posição dos gnetófitos não é consensual; a figura ilustra uma das duas hipóteses com maior apoio molecular — a hipótese das gnepinas (gnepine hypothesis). N.b.: o ancestral comum das angiospérmicas e das acrogimnospérmicas é anterior à evolução das angiospérmicas; as angiospérmicas são substancialmente mais recentes do que as acrogimnospérmicas. [Filogenia baseada em Ran et al. (2018)].
As plantas com semente atuais herdaram de uma gimnospérmica ancestral desconhecida características essenciais do seu ciclo de vida, entre as quais se destacam: (i) o ciclo haplodiplonte com dominância acrescida do esporófito, (ii) a heterosporia, (iii) os gametófitos unissexuais profundamente simplificados, (iv) a retenção do esporo ♀ (endosporia) e (v) do gametófito ♀ (endoprotalia) no interior de um primórdio seminal, (vi) a formação do pólen, (vii) a independência da fertilização em relação à presença de água líquida no exterior e, claro, (viii) a produção da semente.
Recordo que a reprodução sexual das plantas com semente envolve sete processos essenciais resumidos no Quadro 55. Vejamos agora o que mais distingue e aproxima as gimnospérmicas e as angiospérmicas ao nível reprodutivo.
Quadro 55. Processos essenciais da reprodução sexual nas plantas com semente (gimnospérmicas + angiospérmicas)
| Etapa | Descrição |
| Esporogénese | Diferenciação de micrósporos e megásporos no interior de sacos polínicos (microsporângios) e de primórdios seminais (megasporângios), respetivamente. |
| Gametogénese | Desenvolvimento dos gametófitos ♂ (pólen) e ♀ (saco embrionário) a partir, respetivamente, dos micrósporos e dos megásporos. |
| Polinização* | Transporte do pólen (gametófito ♂) entre os sacos polínicos e o micrópilo (nas gimnospérmicas) ou o estigma (nas angiospérmicas). |
| Germinação do grão de pólen e desenvolvimento do tubo polínico* | Envolve a hidratação do pólen e a emergência do tubo polínico. O tubo polínico penetra ativamente no nucelo nas gimnospérmicas, e nos tecidos do estilete nas angiospérmicas. |
| Fecundação (= fertilização) | Inclui a libertação dos gâmetas ♂ pelo tubo polínico e a fusão gamética. |
| Embriogénese | Desenvolvimento do embrião. |
| Formação da semente* | Envolve a diferenciação de um embrião, a acumulação de reservas em tecidos especializados e a formação de um tegumento protetor, culminando este processo com a entrada num estado de quiescência ou dormência. |
* Processo ausente nas plantas de 'esporulação livre'.
As plantas com semente são invariavelmente heterospóricas. Os micrósporos diferenciam-se em grande número no interior de sacos polínicos, interpretáveis evolutivamente como microsporângios. Os megásporos formam-se no interior de primórdios seminais. O megasporângio corresponde à massa celular diploide de origem materna que enche o primórdio seminal, i.e., ao nucelo (v. «Primórdio seminal») — sendo os tegumentos aquisições evolutivas posteriores. O megasporângio das plantas com semente é carnudo e diferencia um único esporo viável (megásporo) indeiscente (o que garante a retenção do megásporo). Nos briófitos e nos 'pteridófitos', os esporângios têm de uma a poucas camadas celulares de espessura e diferenciam inúmeros esporos que são libertados para o exterior na maturação (deiscência). Os primórdios seminais são externamente cingidos por 1 tegumento nas gimnospérmicas ou 1 a 2 tegumentos nas angiospérmicas. O primórdio seminal deve, então, ser entendido anatómica e evolutivamente como um megasporângio tegumentado. A maior parte da bibliografia generalista é menos precisa e confunde o megasporângio com o primórdio seminal; segue-se esta aproximação rigorosa para não complexificar excessivamente o texto.
Em vez dos briófitos e dos 'pteridófitos', nas plantas com semente ocorre uma retenção dos esporos, tanto ♂ como ♀, no corpo do esporófito, i.e., os esporos germinam retidos no interior dos esporângios parentais. Os esporos ♂ dão origem aos grãos de pólen e os ♀ ao saco embrionário. Um grão de pólen unicelular é homólogo a um micrósporo; o pólen pluricelular (germinado/maduro) corresponde ao gametófito ♂, sendo homólogo do microprotalo dos 'pteridófitos' heterospóricos. O saco embrionário — o gametófito ♀ — é homólogo, supõe-se, do megaprotalo dos 'pteridófitos'heterospóricos.
Os gametófitos das ‘plantas de esporulação livre’ têm uma vida terrestre autónoma, ainda que, por vezes, dependente de associações simbióticas com fungos. Nas plantas com semente, a sua diferenciação e manutenção dependem integralmente da geração esporofítica: antes da deiscência, o gametófito ♂ é sustentado pelos tecidos esporofíticos do saco polínico paternal; depois da germinação estigmática, na forma de tubo polínico, é alimentado pelo pistilo (ou nucelo, nas gimnospérmicas) da planta polinizada. O gametófito ♀ é alimentado permanentemente pelo nucelo. Os gametófitos dos espermatófitos, sobretudo nas angiospérmicas, são substancialmente mais simples do que os dos briófitos e dos 'pteridófitos'. Nos espermatófitos, apenas o gametófito ♂ é móvel; o gametófito ♀ não tem vida livre, permanecendo fechado no interior do primórdio seminal, circundado por tecidos esporofíticos maternos (nucelo e tegumentos).
O gametófito ♂ maduro das gimnospérmicas tem 3 ou mais células; o das angiospérmicas tem sempre 3 células. O gametófito ♀ das gimnospérmicas contém vários milhares de células (Linkies et al., 2010), diferenciando vários arquegónios (cada um com uma oosfera inclusa), embora geralmente se forme apenas um embrião viável. O gametófito ♀ das angiospérmicas está drasticamente reduzido a (3-6)7(8) células. Hoje em dia, a biologia evolutiva evita tentar identificar anterídios e arquegónios nos gametófitos das angiospérmicas e nos grupos mais avançados de gimnospérmicas (como haviam ensaiado muitos autores clássicos), porque estes órgãos foram profundamente simplificados ou eliminados pela evolução.
Nas plantas com semente, os primórdios seminais (megasporângios + 1 ou 2 tegumentos) e os sacos polínicos (microsporângios) inserem-se em órgãos de suporte (bearing organs) de origem foliar (esporofilos; sporophylls) ou caulinar (ramos férteis; fertile shoots). Os sacos polínicos desenvolvem-se exclusivamente em órgãos de suporte de natureza foliar: os microsporofilos. Reconhecem-se dois tipos de microsporofilos fundamentais: as escamas polínicas nas gimnospérmicas e os estames nas angiospérmicas. Os órgãos de suporte ♀ são bem mais difíceis de interpretar revolutivamente. Nas gimnospérmicas atuais, consoante o grupo taxonómico, podem ser de natureza foliar (nem sempre assimiláveis ao conceito de megasporofilo típico) ou caulinar (ramos férteis). O carpelo das angiospérmicas, por outro lado, é o paradigma do megasporofilo. Na maioria das angiospérmicas, os órgãos de suporte e os esporângios estão concentrados numa estrutura bissexuada altamente especializada — a flor. As gimnospérmicas não têm verdadeiras flores, possuindo somente estruturas reprodutivas que são invariavelmente unissexuadas.
Após a polinização, as gimnospérmicas mais avançadas ('coníferas' e Gnetidae) e todas as angiospérmicas produzem um tubo polínico pluricelular que conduz os gâmetas ♂, i.e., as células espermáticas, ao encontro da célula gamética ♀, a oosfera. Este processo de fecundação designa-se por sifonogamia. Contudo, os grupos mais antigos de gimnospérmicas — Ginkgoidae e Cycadidae — seguem um modelo de reprodução intermédio entre a oogamia e a sifonogamia, designado por zooidogamia (ou sifonogamia imperfeita): produzem um tubo polínico de função puramente haustorial que rebenta na vizinhança dos arquegónios, libertando gâmetas ♂ flagelados (anterozoides) que são autónomos na sua curta deslocação a nado até à oosfera. Supõe-se que a sifonogamia clássica evoluiu de forma independente nas gimnospérmicas mais avançadas e nas angiospérmicas a partir de um ancestral comum zooidogâmico.
Nas gimnospérmicas, o tubo polínico cresce de forma invasiva no tecido nucelar e geralmente é ramificado. Desempenha duas funções: transportar os gâmetas (função de transporte) e extrair ativamente nutrientes do primórdio seminal em favor do gametófito ♂ (função haustorial). O tubo polínico das angiospérmicas, pelo contrário, não se ramifica, progride muito rapidamente através dos tecidos do estilete e é de muito mais curta duração (vd. «Fase progâmica»). Até à fecundação, o gametófito ♂ é alimentado pelo nucelo do primórdio seminal nas gimnospérmicas, e pelos tecidos esporofíticos do estilete do indivíduo polinizado nas angiospérmicas.
A fecundação determina a síntese do novo esporófito. Nos espermatófitos, o esporófito recém-diferenciado, i.e., o embrião, desenvolve-se encapsulado no primórdio seminal fecundado, sendo alimentado pelo esporófito materno até à maturação da semente. As reservas nutritivas injetadas pelo esporófito materno na semente garantirão a sobrevivência do embrião até que este, depois da germinação e da emergência fotossintética, dê origem a uma plântula autotrófica autónoma. A semente é, assim, um esporófito imaturo num estado quiescente/dormente, provido de tecidos de reserva e envolvido por uma estrutura protetora (o tegumento) com origem nos tegumentos do primórdio seminal materno. Como veremos, as sementes das gimnospérmicas e das angiospérmicas apresentam várias diferenças anatómicas, sendo a mais relevante a origem e a ploidia do tecido nutritivo principal: o próprio gametófito ♀ haploide (frequentemente designado na literatura por endosperma primário) nas gimnospérmicas, em oposição ao verdadeiro endosperma secundário triploide nas angiospérmicas.
Os espermatófitos dispersam-se no ambiente em dois momentos muito precisos do seu ciclo de vida: enquanto pólen (mobilidade do gametófito ♂) ou enquanto semente (mobilidade do esporófito imaturo). Os grupos mais antigos de plantas terrestres (briófitos e 'pteridófitos') deslocam-se sob a forma de esporos (mobilidade espórica) e, de forma espacialmente muito limitada, sob a forma de gâmetas ♂ (mobilidade gamética).
As gimnospérmicas são genericamente anemófilas (polinizadas pelo vento) e anemocóricas (sementes dispersas pelo vento). As interações mutualistas com animais são, nestas plantas, secundárias e residuais. Por oposição, a assombrosa diversidade das relações mutualistas com os animais constitui um dos maiores trunfos evolutivos das angiospérmicas.