
Organização do corpo das plantas com semente
Órgãos fundamentais
A homogeneidade do ambiente aquático ancestral foi pouco propícia à especialização celular e morfológica das plantas (Niklas, 2016). Na terra firma, porém, os recursos encontram-se segregados em dois compartimentos distintos: o solo (água e nutrientes) e a atmosfera (luz e CO₂). Este facto forçou a evolução de estruturas especializadas e polarizadas no corpo das plantas terrestres de modo a poderem «habitar», simultaneamente, a terra e o ar.
O esporófito – a fase diploide do ciclo de vida das plantas terrestres – das plantas vasculares é constituído por três órgãos fundamentais: raiz (root), caule (stem) e folha (leaf)(Troll 1948 cit. Classen-Bockoff 2001; Figuras I.1.4 e I.1.5). Na planta vascular arquétipo, as raízes absorvem água e nutrientes e ancoram o corpo da planta ao solo; os caules providenciam suporte mecânico e o transporte de substâncias entre a raiz e a parte aérea, e vice-versa; as folhas sequestram carbono da atmosfera sob a forma de moléculas carbonadas complexas com a intervenção da luz solar. Todas as estruturas das plantas resultam de modificações durante o processo evolutivo (mudanças evolutivas; evolutionary changes) ou ao longo do ciclo de vida (mudanças ontogénicas; ontogenetic changes) de um destes três órgãos. A flor, por exemplo, é um ramo curto muito modificado, com folhas especializadas na reprodução sexuada.
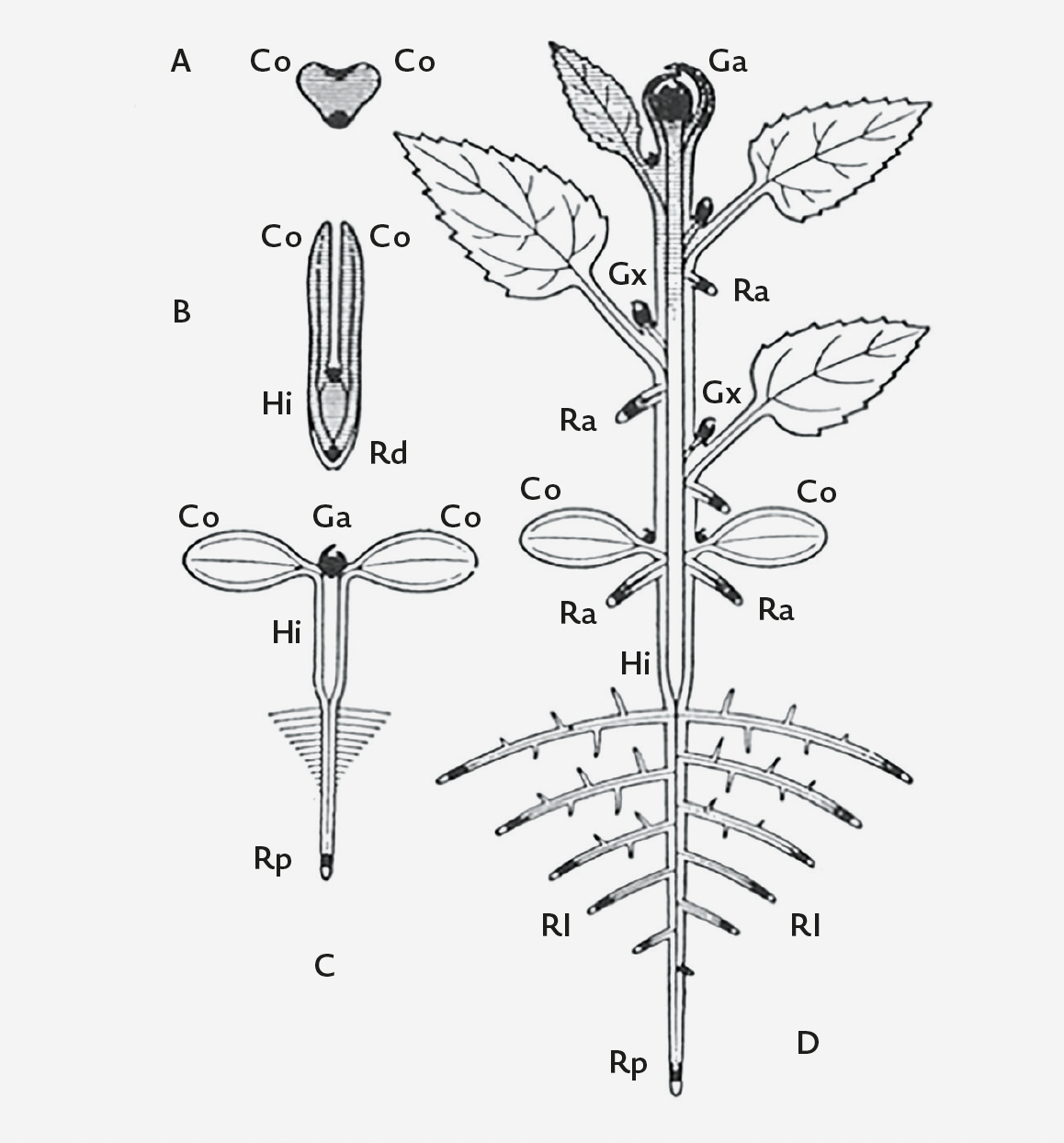
Figura I.1.4. Estrutura do embrião (cormo embrionário) e do cormo pós-embrionário de uma ‘dicotiledónea’. A) Embrião jovem: Co – cotilédone. B) Embrião maduro de uma semente cotiledonar: Hi – hipocótilo; Ra – radícula. C) Plântula: Rd – raiz primária. D) Estrutura do cormo de uma angiospérmica: Ga – gema apical; Rl – raízes laterais; Rd – raízes adventícias; Gx – gemas axilares. [Figura clássica de Sachs adaptada por Troll (1954).]

Figura I.1.5. Estrutura do cormo. Plântula de Lupinus albus (Fabaceae); comparar com a Figura I.1.4. [Fotografia do autor.]
O conjunto das raízes, caules e folhas constitui o cormo (corm) ou corpo da planta (plant body). As partes do cormo não envolvidas na reprodução sexuada compõem o sistema (ou corpo) vegetativo. O sistema (ou corpo) reprodutivo compreende as estruturas reprodutivas nas gimnospérmicas, ou as inflorescências e as flores nas angiospérmicas.
As raízes e os caules exibem a nível anatómico uma simetria radial e, geralmente, têm um crescimento indeterminado (indeterminate growth) — i.e., mantêm meristemas ativos capazes de produzir novos tecidos indefinidamente. As folhas, pelo contrário, são, salvo raras exceções, determinadas e de simetria bilateral (v. «Simetria»). Ao contrário dos órgãos determinados, nos órgãos de crescimento determinado (determinate growth; «v. Alongamento rameal»), como sejam as folhas e alguns tipos de caules (e.g., esporões), o crescimento e a diferenciação estão sujeitos a um estreito controlo genético, pouco sensível a fatores ambientais. As folhas representam um caso extremo de determinação porque, concluída a sua diferenciação, mantêm a mesma forma e estrutura interna até à senescência. Estão descritas algumas, muito raras, exceções. Existem, contudo, raras exceções à determinação foliar: as folhas têm crescimento indeterminado (basal contínuo) na icónica Welwitschia mirabilis (Figura I.1.6) e em vários membros da família Gesneriaceae.

FIGURA I.1.6. Folhas de crescimento indeterminado. A Welwitschia mirabilis (Welwitschiaceae, Gnetidae) produz dois, eventualmente três, pares de folhas (von Willert 1993). O primeiro par – os cotilédones – é substituído por duas folhas opostas longas (até 4 m) persistentes durante toda a vida da planta (von Willert 1995). A produção de novas folhas cessa em seguida, ficando a planta truncada na extremidade distal. As duas folhas alongam-se continuamente pela base a partir de meristemas intercalares foliares situados na interface da folha com a coroa. A destruição dos meristemas intercalares pela herbivoria compromete irremediavelmente a sobrevivência da planta. [Angola, Deserto do Namibe; fotografia do autor.]

FIGURA I.1.7. Caules articulados de Casuarina equisetifolia (Casuarinaceae). N.b.: folhas verticiladas escamiformes, muito curtas. [Cortesia de Paulo Araújo.]
As raízes inserem-se, geralmente, no colo (região de encontro do caule com o sistema radicular) ou em outras raízes. As raízes adventícias (adventitious roots), por definição, são emitidas por raízes lenhosas ou inserem-se em qualquer ponto da parte aérea, sobretudo nos nós dos caules, imediatamente abaixo da inserção das folhas (v. «Tipos de raízes»). As raízes distinguem-se dos caules por possuírem uma coifa (ou caliptra) na extremidade e pela ausência de nós e entrenós. As raízes jamais possuem folhas, embora, por vezes, possam diferenciar gemas adventícias que, mais tarde, dão origem a novos caules e folhas. Os caules de origem radicular são designados nas plantas lenhosas por pôlas, rebentões radiculares ou drageões, e nas plantas herbáceas vivazes por rebentos radiculares
As folhas inserem-se, num padrão regular, obliquamente nos nós (= verticilos caulinares; node) (Figuras I.1.4-D e I.1.5). O entrenó (internode) corresponde à porção de caule entre dois nós sucessivos. Nas éfedras (Ephedraceae) e nas casuarinas (Casuarinaceae) os entrenós destacam-se com facilidade — são articulados (Figura I.1.7). Na axila de cada folha encontra-se, geralmente, pelo menos uma gema (v. «Gemas»; bud), i.e., um aglomerado de células indiferenciadas com capacidade de se multiplicarem indefinidamente (v. «Meristemas»), envolvido por esboços de folhas, complementado, ou não, por um revestimento externo de folhas de proteção escamiformes (catafilos; «v. Tipos de filomas»). A queda das folhas deixa uma cicatriz folhear[1] (leaf scar) no nó, cuja forma tem valor diagnóstico em algumas famílias de plantas com flor (e.g., Moraceae e Juglandaceae; Figura 215-E).
Na extremidade distal dos ramos, situam-se uma gema apical (apical bud) e, no seu interior, um meristema apical caulinar. As gemas axilares (axilary buds) e os respetivos meristemas axilares localizam-se, como se depreende do termo, na axila das folhas (Figuras I.1.4-D e I.1.5). Os meristemas apicais alongam os caules; os meristemas axilares ramificam-nos.
Nas plantas com semente, a formação do cormo inicia-se com a germinação da semente e, implicitamente, com a retoma do crescimento do embrião. O cormo embrionário consta de (Figuras I.1.4-B):
- Radícula (raiz embrionária; radicle) – esboço de raiz;
- Cotilédones (cotyledon) – filomas embrionários ricos em reservas ou com uma função haustorial (v. «Embrião»);
- Plúmula ou gémula (plumule) – esboço de caule com folhas embrionárias envolvendo o meristema apical embrionário.
Nas eudicotiledóneas, os entrenós situados abaixo e acima dos cotilédones são conhecidos, respetivamente, por hipocótilo (hypocotyl) e o epicótilo (epicotyl). Nas Poaceae, o primeiro entrenó do caule acima do escutelo[2] (v. «Semente e germinação do milho-graúdo») merece uma designação especial: mesocótilo (mesocotyl). O colo corresponde à zona de transição entre a raiz e o caule. Na semente, como se verá, o embrião está acompanhado por tecidos de reserva, e o conjunto «embrião + tecidos de reserva» protegido por um tegumento.
O corpo de plantas e animais: estudo comparativo
A evolução das plantas (Archaeplastida) ocorreu numa etapa precoce da história evolutiva dos eucariotas. Os estudos filogenómicos suportam a hipótese da origem dulçaquícola da endossimbiose fundadora da linhagem dos eucariotas fotossintéticos e situam a sua evolução na primeira metade do Paleoproterozoico, há ~2100 Ma; o ancestral comum das plantas atuais, i.e., o grupo-coroa das arqueplastidas, terá ~1900 Ma (Sánchez-Baracaldo et al., 2017)[3]. Por conseguinte, plantas e animais atuais evoluem independentemente há mais de 2000 Ma, consolidando a sua separação no momento em que a cianobactéria ancestral de todos os cloroplastos ficou retida e se reproduziu no interior da alga primordial. Enquanto os animais prosseguiram na senda da heterotrofia herdada do último ancestral comum aos dois clados, as plantas exploraram as oportunidades evolutivas abertas pela autotrofia. Estão em causa duas formas antitéticas de obter energia. Não surpreende, por isso, que a ontogénese e a estrutura do corpo dos animais e das plantas sejam tão diferentes.
Exploremos o que mais os distingue.
Fotoautotrofia vs. heterotrofia
As plantas, tal como todos os seres vivos que povoam o planeta Terra, são máquinas biológicas e, como tal, estão sujeitas à segunda lei da termodinâmica: para crescerem, reproduzirem-se ou, simplesmente, para permanecerem vivas, consomem e dissipam energia. Uma interrupção no abastecimento de energia traduz-se numa desorganização das células e na morte dos indivíduos. Os animais são heterotróficos: obtêm a energia de que necessitam e constroem o seu corpo a partir de compostos orgânicos complexos adquiridos por ingestão. As plantas têm um metabolismo energético distinto – são seres fotoautotróficos[4]: pepela intervenção da molécula verde, a clorofila, no processo da fotossíntese, sequestram a energia solar sob a forma de energia química (ATP) e poder redutor (NADPH). As plantas gastam menos energia por unidade de biomassa para se manterem vivas do que os animais.
As plantas têm um anabolismo completo: sintetizsintetizam 100% dos seus aminoácidos, proteínas, lípidos, ácidos nucleicos, vitaminas e outras biomoléculas a partir da energia química e do poder redutor disponibilizados pela fotossíntese, mais a água, CO₂ e 20 substâncias minerais simples (e.g., K⁺ e NO₃⁻) absorvidas pelas raízes (Wise & Hoober cit. Crang et al. 2018). Os animais dependem de um vasto número de substâncias extraídas do corpo dos vegetais ou animais que lhes servem de alimento, ou dos resíduos por eles produzidos. Por exemplo, o corpo humano sintetiza apenas 11 dos 20 aminoácidos proteinogénicos (que fazem parte das proteínas), estando todos eles disponíveis nas plantas.
Embora seja a fonte primária de toda a matéria orgânica e energia que percorre os ecossistemas terrestres, a fotossíntese é um processo de conversão energética pouco eficiente. O máximo teórico da eficiência de conversão da energia solar incidente total em energia armazenada sob a forma de biomassa é de 4,6% nas plantas C3 e 6% nas plantas C4 (Zhu et al., 2008). Este valor é substancialmente mais baixo no campo porque as plantas jamais crescem em condições ecológicas ótimas. Em contrapartida, a eficiência da conversão da energia solar total em energia elétrica em painéis solares fotovoltaicos experimentais atingiu os 47,6%, e ronda os 17-20% nos modelos comerciais (Schygulla et al., 2022). Os livros de bioquímica dizem-nos que a oxidação da glucose – o processo fundamental de produção catabólica de energia química da célula eucariota – é bem mais eficiente: cerca de 40%, um valor superior a um motor a combustão (ca. de 30%). As plantas necessitam de grandes superfícies de captação e conversão de energia luminosa expostas ao sol para satisfazerem as suas necessidades energéticas; os animais prosperam com concentrados de energia química.
Transporte de fluídos aquosos e mobilidade celular
A água é o constituinte maioritário de todos os seres vivos. Porém, o papel da água na fisiologia das plantas não tem equivalente animal. Por não terem tecidos musculares capazes de gerar força, as plantas evoluíram de modo a tirar partido das propriedades físicas da água para realizar trabalho. A transpiração resultante da diferença entre a energia da água do solo e da atmosfera é suficiente para pôr em marcha um sistema circulatório, e elevar a água pelo xilema mais de 100 m acima do solo numa Sequoia. Mais de 95% da água transportada pelo xilema regressa à atmosfera pelos estomas; as plantas terrestres contribuem, anualmente, com cerca de 10% do vapor de água da atmosfera (Hanson, 1991). A força gerada pela concentração diferencial de solutos através de membranas semipermeáveis – força osmótica – propulsiona a seiva floémica. Quando combinada com o relaxamento diferencial por via enzimática da parede celular controla a expansão celular, o crescimento e várias formas de movimento (Cosgrove, 2016). Enquanto os animais usam os músculos para bombear, puxar e empurrar, as plantas servem-se da água para bombear, empurrar e expandir células vivas e órgãos (Crang et al., 2018).
A migração celular a longa distância no interior do corpo é uma característica animal. O sangue, por exemplo, transporta vários tipos celulares, enquanto a seiva das plantas é uma solução aquosa diluída acelular. Os anterozoides das ‘plantas de esporulação livre’, do ginkgo e das cicas, e as células constituintes do tubo polínico das restantes gimnospérmicas e das angiospérmicas são as únicas exceções vegetais conhecidas (v. «Fase progâmica»). A evolução da parede celular explica, pelo menos em parte, a imobilidade das células vegetais.
Esta imobilidade faz com que a orientação das divisões celulares a nível meristemático estabeleça a posição espacial das células-filhas no contexto dos órgãos em desenvolvimento. A posição das células, por sua vez, condiciona a sua diferenciação. As células vegetais são capazes de «percecionar» a sua posição relativa nos meristemas e, em face disso, modular o perfil de expressão genética e diferenciar-se num tipo celular adequado (Kwak et al., 2005; Singh & Bhalla, 2006).
Perceção das condições ambientais e comunicação
Os seres vivos estão expostos a inúmeros fatores de stresse ambiental como a presença de competidores, de parasitas, de herbívoros ou de predadores, a condições extremas de secura, calor e frio, ou a desequilíbrios nutricionais no solo ou nos alimentos ingeridos. A reação e a sobrevivência ao stresse ambiental dependem, necessariamente, da perceção do ambiente exterior e da comunicação interna entre as partes (e.g., entre órgãos).
Os sistemas de perceção ambiental das plantas são extraordinariamente diversos e sofisticados. As plantas são sensíveis a sinais físicos (e.g., às alterações da qualidade da luz, temperatura, ao toque e ao consumo de partes) e químicos (e.g., disponibilidade de nutrientes no solo, substâncias químicas libertadas por parasitas ou pelos caules e raízes das plantas vizinhas). A comunicação interna da leitura do ambiente exterior envolve, sobretudo, a emissão de sinais químicos que circulam pelo floema; e.g. aumento súbito da disponibilidade de fotoassimilados, fito-hormonas, mRNA e vários tipos de RNA não codificante (Kehr & Kragler, 2018). Certos mecanismos de comunicação têm características análogas aos impulsos nervosos animais; e.g., sinais de glutamato transmitidos ao longo do floema em resposta ao ataque de fitófagos (Toyota et al., 2018) ou os impulsos elétricos ao longo dos feixes vasculares nas folhas sujeitas a pressão mecânica (T. H. Yang et al., 2023). evidência sugere até que a resposta das plantas a sinais ambientais é condicionada por experiências passadas, um fenómeno designado por priming (preparação) (Calvo et al., 2020). A Através de alterações epigenéticas (modificações na cromatina que não alteram a sequência de ADN), uma planta que sofreu um ataque prévio consegue reagir mais rápida e vigorosamente a um novo ataque, demonstrando uma forma de "memória" bioquímica sem necessidade de armazenamento neural (Lin et al., 2022). Pese embora a natureza descentralizada do corpo das plantas, só assim se explica que os estragos provocados por insetos desencadeiem respostas por antecipação em outras folhas, ou que o crescimento de uma árvore após a perda de partes se faça de forma equilibrada.
A comunicação entre indivíduos é igualmente adaptativa. Foi provado que as plantas comunicam entre si, através de exsudados radiculares, reconhecendo o grau de parentesco genético da vizinhança, e que essa comunicação e que essa comunicação influencia o crescimento e a forma (Chen et al., 2012). As plantas emitem também compostos voláteis de aviso ao nível da canópia perante um ataque de parasitas e herbívoros (Ueda et al., 2012) e as raízes de indivíduos compatíveis enxertam-se umas nas outras trocando todo o tipo de moléculas orgânicas (Lev-Yadun & Sprugel, 2011). Para além do contacto físico direto, a comunicação subterrânea é amplificada pelas redes micorrízicas comuns (a chamada Wood Wide Web). Os fungos simbiontes, cujas hifas conectam as raízes de múltiplos indivíduos, funcionam como "autoestradas" biológicas, permitindo a transferência de carbono, azoto e sinais de alerta químicos entre árvores vizinhas, mesmo de espécies diferentes (Simard, 2018). Embora a interpretação destes fluxos como atos de "altruísmo" seja contestada, a existência da rede física de partilha é um facto ecológico. Estudos recentes mostram que as plantas, além dos compostos voláteis, são capazes de emitir ultrassons relacionados com a hidratação celular ou o ataque de insetos fitófagos, específicos para cada espécie vegetal e tipo de stresse, sendo expectável que sejam detetados por indivíduos coespecíficos ou não (Khait et al., 2023). A comunicação entre as plantas é um tema de ponta da fisiologia vegetal.
A perceção, a comunicação e a resposta a sinais ambientais é um pressuposto para persistência da vida num planeta em constante mudança. Embora o propósito seja o mesmo, as variáveis ambientais percecionadas, e os mecanismos e órgãos envolvidos, a informação extraída e as respostas são distintos nos animais e nas plantas. As plantas não têm paladar nem audição e os animais não sentem a falta de nutrientes minerais no solo. O fitocromo das plantas e o olho dos vertebrados são sensíveis à luz mas evoluíram de forma independente: não geram a mesma informação e muito menos as mesmas respostas. As plantas não têm um sistema nervoso como os animais e um cérebro a centralizar o processamento da informação sensorial. A biologia «sensorial» de animais e plantas não é homóloga. O «mundo sensível» das plantas e dos animais têm pouco em comum.
Por falta de um vocabulário adequado, a descrição dos «sentidos» das plantas é frequentemente feita com termos e conceitos importados da zoologia ou da psicologia humana, um facto que tem dado origem a sérios equívocos. É mais correto dizer que as plantas percecionam e respondem a estímulos exteriores, alguns deles sentidos, mas de outra maneira, pelos animais; e.g., temperatura, humidade do ar, comprimento do dia, intensidade da luz. Defender que as plantas conversam com outros organismos e entre si usando os compostos orgânicos voláteis como o seu alfabeto, que são inteligentes ou que têm sentimentos (e.g., medo, compaixão) são abusos grosseiros da metáfora (Calvo et al., 2020; Loreto & D’Auria, 2022; Wohlleben, 2017).
A inteligência vegetal – um tema na moda em alguns meios ligados à botânica – é definida do seguinte modo: «qualquer tipo de comportamento intencional e flexível que seja benéfico e permita ao organismo atingir o seu objetivo» (Calvo et al., 2020). Algumas bactérias detetam a iminência de um stresse ambiental pela intervenção de proteínas sensíveis à luz (Hatfield et al., 2023) – são também inteligentes? A inteligência é um atributo animal, conforme se depreende numa das definições mais citadas da psicologia – «a inteligência é uma capacidade mental muito geral que, entre outras coisas, envolve a capacidade de raciocinar, planear, resolver problemas, pensar abstratamente, compreender ideias complexas, aprender rapidamente e aprender com a experiência» (Gottfredson, 1997). Pela repetição arbitrária, a metáfora ganha novos significados inadequados para a descrição e compreensão da realidade.
Volume e superfície nas plantas
A química da vida é essencialmente uma química do carbono. O CO2, o nutriente carbonado das plantas, é absorvido do ar pelos caules primários e pelas folhas. O ar é uma solução gasosa à base de azoto, com uma concentração muito baixa de CO2: 0,042% em volume (420 ppm). Como se referiu antes, a fotossíntese é intrinsecamente ineficiente. A otimização evolutiva da absorção da luz e do CO2 fez-se através do aumento da superfície em detrimento do volume da parte aérea (maximização da relação superfície/volume). Portanto, as folhas laminares são uma adaptação à captura de luz e à assimilação de CO2.
Os restantes nutrientes que compõem a massa das plantas são maioritariamente absorvidos pelas raízes de uma solução igualmente muito diluída – a água do solo –, ou extraídos da superfície dos minerais e da matéria orgânica do solo. O sistema radicular foi sujeito a pressões de seleção análogas. A captura de nutrientes, sobretudo de baixa mobilidade, como o fósforo, depende da exploração de um grande volume de solo por uma extensa rede de raízes cilíndricas. Neste caso, o aumento da relação superfície/volume fez-se à custa da minimização do diâmetro das raízes.
Num trabalho clássico de quase um século, com plantas de centeio (Secale cereale) cultivadas durante 4 meses numa caixa de solo de ca. 0,1 m3, Dittmer (1937) calculou que o comprimento total das raízes atingia 622,8 km e o dos pelos radiculares 10.628 km, com uma superfície total de 639 m², contribuindo os pelos radiculares com cerca de 400 m². Hallé (2002) estimou que a superfície externa da parte aérea (caules + folhas) de uma árvore com 40 m de altura possa ultrapassar 1 ha. A superfície das raízes é ainda maior. A relação superfície da parte aérea/superfície da parte subterrânea é muito variável; Hallé (2002) refere um valor meramente indicativo de 1:130. Assim sendo, a superfície externa das raízes dessa árvore de 40 m pode atingir os 130 ha! Este valor é substancialmente maior se for contabilizada a superfície das hifas dos fungos micorrízicos (v. «Micorrizas»).
Os animais como seres unitários
Os animais (metazoários) são, genericamente, organismos unitários (= não modulares). Esta classificação fundamenta-se em três razões: (i) ao contrário das plantas, o corpo animal não resulta da acumulação reiterada de unidades multicelulares discretas (módulos); (ii) as partes em que se divide o corpo dos animais unitários estão organizadas hierarquicamente e funcionalmente integradas; e (iii) os animais exibem um crescimento e uma estrutura determinados.
O produto da embriogénese animal é, em essência, uma versão em escala reduzida de um indivíduo adulto, com a organogénese já iniciada. As estruturas externa e interna dos animais unitários – e.g., o sistema vascular, o número de membros e a posição espacial do aparelho digestivo ou dos pulmões de um mamífero ou de uma ave – mantêm-se praticamente inalteradas durante o crescimento pós-embrionário (Figura 1.8). Adicionalmente, o volume e a massa do corpo estabilizam ao atingir o estado adulto. Mesmo os insetos holometábolos[5], apesar de sofrerem modificações estruturais profundas durante o desenvolvimento, transitam de forma determinada entre os estádios de ovo, larva, pupa (ou crisálida) e adulto, sendo a morfologia final (o imago) definitiva. Nos animais adultos, as células estaminais têm a função primária de repor células especializadas de duração limitada (e.g., células sanguíneas ou epidérmicas) (Baürle & Laux, 2003). Em contraste, como se verá, as plantas possuem um corpo morfologicamente flexível onde as células estaminais geram continuamente novos órgãos.
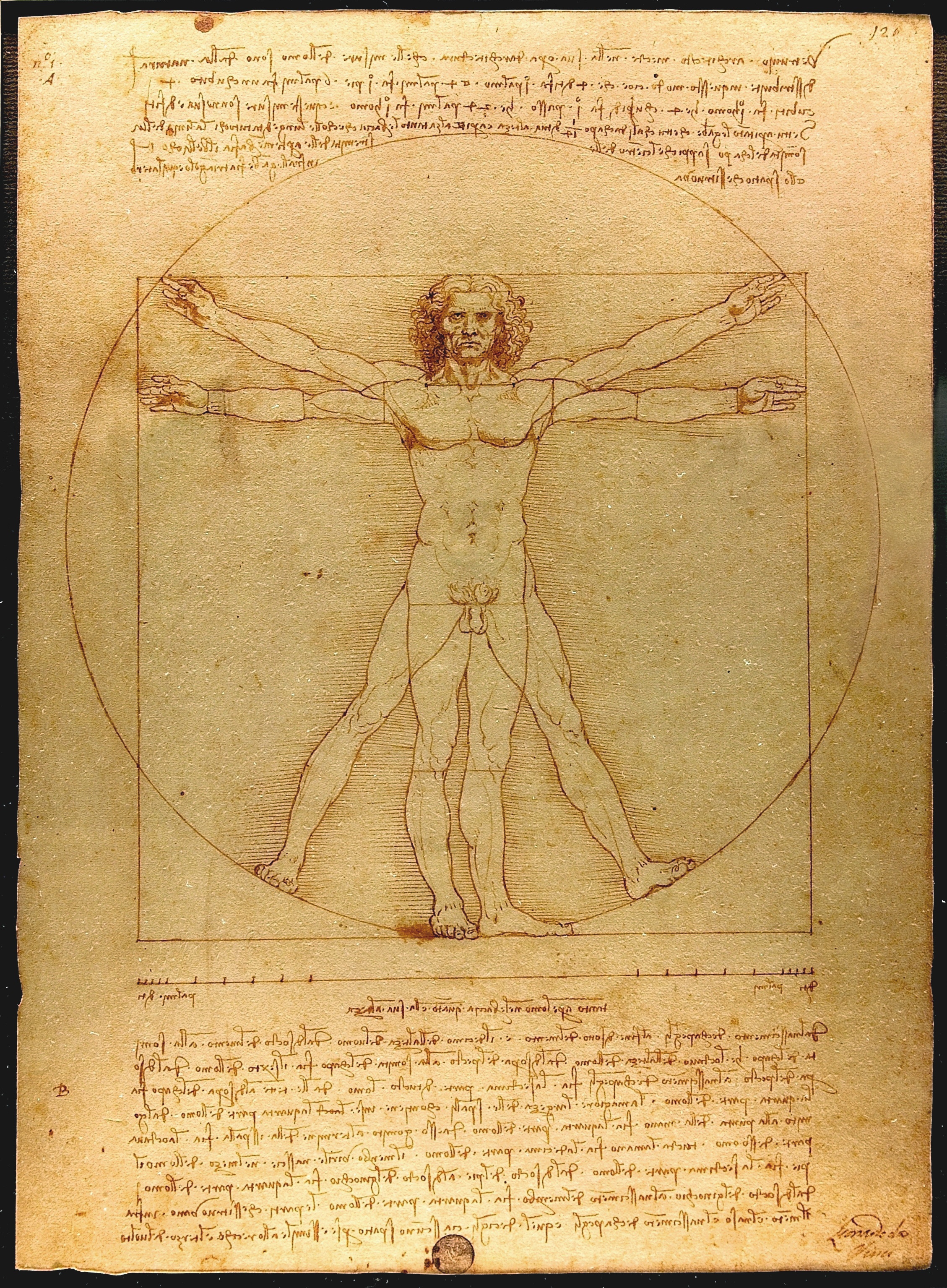
FIGURA I.1.8. O corpo animal. As partes em que se divide o corpo dos animais unitários estão organizadas hierarquicamente, são interdependentes e têm, aproximadamente, a mesma idade. As estruturas externa e interna encontram-se sob forte controlo genético, mantendo-se praticamente inalteradas durante o crescimento pós‑embrionário. Genericamente, os seres humanos adultos são mais semelhantes entre si do que duas plantas coespecíficas. O Homem Vitruviano de Leonardo da Vinci (ca. 1490) é uma representação fidedigna das proporções da forma humana; uma tarefa difícil de replicar com o corpo variável de muitas plantas. [Wikimedia Commons.]
As partes constituintes do corpo dos animais unitários estão organizadas hierarquicamente, são interdependentes e possuem, aproximadamente, a mesma idade, ainda que coexistam células em diferentes estádios de renovação. Esta estrutura é hierarquizada por ser constituída por um elevado número de tipos celulares especializados – no Ser Humano, cerca de 400 grandes tipos celulares distribuídos por 60 tipos de tecidos (Hatton et al., 2023) –, os quais se organizam, por sua vez, em órgãos e sistemas com funções definidas e permanentes. A organização hierárquica e a interdependência funcional são tão profundas que os indivíduos necessitam da totalidade, ou da quase totalidade, das suas partes para a manutenção da vida. Um vertebrado não sobrevive sem coração e a falta de um membro locomotor, por exemplo, compromete seriamente o seu sucesso reprodutivo. Esta integração estende-se à morfologia e às relações forma-função, a tal ponto que os paleontólogos de vertebrados conseguem reconstruir esqueletos, corpos e ecologias a partir de fragmentos dispersos de poucos ossos e dentes – um privilégio raro em paleobotânica (Cronquist, 1988).
Existe uma correlação inversa universal entre a especialização funcional das células e a sua totipotência. Por conseguinte, em condições normais, os tipos celulares animais muito especializados são incapazes de reverter à condição de célula estaminal. Este facto ajuda a explicar por que razão a regeneração de partes perdidas por efeito da idade, doença, acidente ou predação (e.g., morte de células neuronais, necrose de tecido cardíaco, ou a perda de membros) é muito limitada nos grupos animais de simetria bilateral. Nas plantas, pelo contrário, a desdiferenciação – a reversão de células diferenciadas para a condição meristemática – é um evento comum (v. «Meristemas»).
A dimensão e as estruturas interna e externa dos seres unitários encontram-se sob um rigoroso controlo do genoma (o plano corporal ou bauplan), sendo, por isso, pouco plásticas face a fatores ambientais. O número e a forma dos órgãos reprodutivos nos animais são determinados numa fase embrionária. Independentemente da disponibilidade de recursos, o número de órgãos e a sua disposição espacial são constantes – nos mamíferos existem apenas um coração, dois rins e quatro membros (dois anteriores e dois posteriores). As plantas, por sua vez, produzem órgãos vegetativos (e.g., filídios, raízes, caules e folhas) e órgãos reprodutivos (e.g., anterídios, cones e flores) em número variável e, frequentemente, de forma escalonada ao longo do tempo. Consequentemente, o corpo dos animais, ao contrário do corpo das plantas, possui uma capacidade muito limitada de ajuste estrutural à disponibilidade de recursos. Este ajustamento ocorre, antes, ao nível da população através de variações na densidade populacional: os animais multiplicam-se com celeridade quando o alimento abunda; na escassez, migram ou sofrem mortalidade em massa.
O movimento nas plantas e animais
Quando o alimento escasseia, os animais não têm outra opção senão mover-se – mudar de lugar. A heterotrofia é indissociável da locomoção. A capacidade de deslocação é igualmente uma resposta-chave dos animais a outros tipos de stresse causados, por exemplo, pela sazonalidade ambiental, pela predação ou pelo parasitismo. Alguns grupos de animais são secundariamente sésseis (evoluíram a partir de ancestrais móveis) durante grande parte do seu ciclo de vida. Para se alimentarem, dependem da deslocação do alimento na sua direção. Os corais (Cnidaria: Octocorallia) e as anémonas (Cnidaria: Hexacorallia) dispõem de tentáculos móveis arranjados em torno de uma abertura oral para capturar as presas que circulam na sua vizinhança.
À exceção de fases muito precisas e breves do seu ciclo de vida, sob a forma de semente ou de pólen (dispersão), a natureza séssil das plantas impede-as de migrar em resposta a um ambiente em permanente mudança. As plantas não possuem membros ou apêndices de locomoção como os animais, nem conseguem desenraizar-se para se fixarem novamente noutro local do solo. Ainda assim movimentam-se, mas de um outro modo e sem mudar de lugar: crescendo.
Designa-se por tropismo o crescimento direcional de órgãos vegetais em resposta a um estímulo externo. Os tipos principais são o fototropismo, o gravitropismo, o tigmotropismo, o hidrotropismo, o oxitropismo e o eletrotropismo, respetivamente, o crescimento em resposta à luz, gravidade, contacto mecânico, humidade, oxigénio e campos elétricos. Os tropismos são mais exuberantes e diversos na raiz do que no caule. Charles Darwin foi pioneiro no estudo dos tropismos vegetais (Charles Darwin, 1880).
O azoto é absorvido pelas plantas em duas formas de grande mobilidade no solo: os iões nitrato e amónio. A maior parte do azoto mineral penetra na planta arrastado pela água absorvida pelas raízes, de forma passiva, através de um mecanismo designado por fluxo de massa. Portanto, as plantas também dependem do movimento de alguns tipos de nutrientes na sua direção, tal como os corais ou as anémonas dependem da passagem das presas. Por oposição, os nutrientes pouco móveis, como o fósforo, têm de ser ativamente procurados pela interceção das raízes em crescimento no interior do solo.
O crescimento é, então, a forma primordial que as plantas têm de se movimentar em busca de recursos. Este crescimento, embora sujeito a interrupções de vária ordem, persiste durante todo o ciclo de vida – é contínuo. As múltiplas funções do crescimento vegetal serão desenvolvidas adiante (v. «Crescimento indeterminado nas plantas»).
Características e consequências da estrutura modular
As plantas são organismos modulares de crescimento indeterminado. Modulares porque constituídos pela repetição de unidades multicelulares discretas, i.e., módulos (modules), de grande autonomia funcional. A semiautonomia dos módulos permite que o crescimento, ao nível do indivíduo, seja matematicamente modelado de forma análoga a uma comunidade de organismos similares e semi-independentes, em competição por recursos escassos, correspondendo cada «organismo» a um módulo individual. De crescimento indeterminado pelo facto de crescerem continuamente até à senescência/morte, ainda que este crescimento possa ser interrompido por períodos de quiescência ou dormência mais ou menos prolongados em condições ambientais desfavoráveis.
O fitómero
A estrutura modular é morfologicamente evidente na parte aérea do corpo das plantas. As raízes, embora cresçam de forma iterativa, não se decompõem em módulos exteriormente distintos (com nós e entrenós) e têm um crescimento oportunista, dirigido por gradientes de oxigénio, água e nutrientes no solo. Os caules, pelo contrário, são construídos por meio do «encaixe» sucessivo, como numa construção de Lego, de um módulo elementar – o fitómero (= metâmero; metamer, phytomer) – constituído por um entrenó, um nó e uma ou mais folhas com os respetivos meristemas axilares (Figura I.1.9)[7] [8]. O fitómero, por sua vez, organiza-se em módulos de complexidade crescente; e.g., pernadas, sistemas de ramos e ramos.

FIGURA I.1.9 Fitómero. A) Fitómero nas monocotiledóneas (numa gramínea): a – fitómero; b – bainha da folha; c – limbo da folha. B) Fitómero nas dicotiledóneas (Jasminum fruticans, Oleaceae). [Fotografias do autor.]
As plantas crescem por meio da adição de módulos construídos por meristemas apicais. Durante o ciclo de vida, dentro dos limites impostos a cada momento pela genética, ajustam o número, a disposição espacial, a forma dos módulos e, por vezes, a sua identidade, às condições ambientais (e.g., temperatura) e à disponibilidade de recursos (e.g., água, luz e nutrientes).
Uma planta de trevo-branco, intensamente desfoliada por mamíferos, adquire folhas pequenas, entrenós curtos e hábito prostrado (Figura I.1.10)[9]. Sob uma secura edáfica extrema, muitas plantas reduzem o número de partes aéreas, rejeitam ramos (v. «Cladoptose») e/ou folhas e, eventualmente, alteram a identidade dos módulos antecipando ou atrasando a produção de flores. O sistema radicular é igualmente deprimido e parcialmente descartável sob stresse (v. «Plasticidade fenotípica do sistema radicular»). Estima-se que 90% das plantas lenhosas são capazes de descartar partes do corpo em condições de stresse extremo, como se refere adiante, a «Setorização hidráulica» tem um papel-chave neste processo (Salguero-Gómez & Casper, 2011).
Por norma, quando as condições ambientais são propícias e os recursos abundantes, os meristemas caulinares ativos são mais numerosos, o número e o comprimento dos módulos aumentam, os caules são mais ramificados e longos, e, atingida a fase reprodutiva, as inflorescências são mais numerosas e com mais flores. Em condições de elevado stresse ambiental, são construídos menos módulos e estes são mais curtos, e as plantas têm um hábito mais congesto e podem rejeitar partes do corpo. As alterações induzidas pelo ambiente na morfologia externa, mas também na fisiologia ou no programa ontogenético, ainda que mediadas pelo aparato genético, caem no conceito de plasticidade fenotípica (phenotypic plasticity)[10].

FIGURA I.1.10. Plasticidade fenotípica dos módulos. Plantas de trevo-branco (Trifolium repens, Fabaceae) não perturbadas (A) e intensamente perturbadas pela herbivoria (B). As plantas sujeitas a desfoliação intensa e contínua têm folíolos pequenos, pecíolos curtos frequentemente peludos e estolhos finos com entrenós curtos. Estas características regridem com a suspensão do pastoreio (Brougham 1958). N.b.: a dimensão das inflorescências e das flores é largamente independente da pressão de pastoreio – não são plásticas como o corpo vegetativo. [Fotografias do autor.]
Proprioceção
Como se referiu no ponto anterior, a organização espacial dos módulos e o crescimento da parte aérea são menos oportunistas do que nas raízes; caso contrário, a copa das árvores seria fortemente assimétrica, orientada para onde vem a luz. Nas latitudes mais elevadas do hemisfério Norte, as árvores tombariam com a idade para sul, e para norte no hemisfério Sul. Como pode, então, uma árvore constituída por milhões de módulos «egoístas» manter uma estrutura estável e equilibrada? A solução deste paradoxo não está totalmente compreendida, mas sabe-se que reside num equilíbrio dinâmico entre a procura de luz (fototropismo), o controlo apical e a capacidade da árvore «sentir», no câmbio, a gravidade e os desequilíbrios mecânicos no interior da canópia (proprioceção, proprioperception) (Moulia et al., 2011).
A árvore responde aos sinais de tensão e compressão detetados pelo câmbio através da formação de lenho de reação. Este crescimento é assimétrico: a planta adiciona lenho nas regiões sob stresse (e.g., na base de um ramo) de modo a colocar (parcialmente) o ramo na posição correta e diminuir o risco de quebra, enquanto contém o seu crescimento em extensão através da redução da adição de módulos (Moulia et al., 2011). Além desta regulação biomecânica, a hierarquia dos ramos é mantida pelo controlo apical: os meristemas apicais localizados na extremidade da árvore (na flecha ou nos ramos principais) enviam sinais hormonais (auxina) para baixo, inibindo a verticalização excessiva dos ramos laterais e garantindo que os módulos «egoístas» se mantêm subordinados à estrutura global e não põem em causa a estabilidade mecânica da árvore.
Setorização hidráulica
Em muitas espécies de plantas, o sistema de transporte não funciona como uma rede de distribuição homogénea de água e solutos, mas sim como uma rede de unidades hidráulicas independentes, com conexões diretas e restritas entre raízes, tronco, pernadas e ramos específicos. Esta segmentação do corpo da planta designa-se setorização hidráulica (hydraulic sectoriality). Este fenómeno explica, em boa parte, a semiautonomia funcional dos módulos vegetais a várias escalas (Salguero-Gómez & Casper, 2011).
Assim, um ramo maduro é interpretado como um setor da planta parcialmente autónomo na produção de fotoassimilados, exportando os excedentes (via floema) para os eixos desprovidos de folhas (troncos e pernadas) e para as raízes. Importa notar que, devido ao transporte no floema ser fortemente axial (vertical) e a difusão lateral limitada, o ramo abastece, preferencialmente, o sistema radicular que se encontra na sua linha vertical direta, bem como a porção do tronco e as pernadas onde se insere.
Esta arquitetura tem implicações práticas: a poda de um ramo de grandes dimensões traduz-se, frequentemente, numa redução do crescimento secundário (em espessura) desse lado específico da árvore, fazendo com que o tronco ou as pernadas adjacentes se tornem assimétricos em corte transversal (Sprugel et al., 1991). A distribuição de fotoassimilados também não é uniforme no sentido vertical. Durante a formação de árvores jovens, a eliminação precoce de ramos laterais baixos (na tentativa de definir demasiado cedo o fuste e a copa definitiva) gera troncos delgados e cilíndricos, frágeis e sensíveis ao vento. Manter estes ramos laterais temporários (ou mesmo "ramos ladrões" controlados) por um ou dois anos é uma boa opção técnica para estimular o espessamento da base e obter troncos precocemente sólidos.
A setorização hidráulica possui um elevado valor adaptativo em ambientes de disponibilidade hídrica limitada ou irregular. Ela circunscreve a propagação de embolias (falhas no xilema) a setores específicos da copa e otimiza o uso de recursos, permitindo à planta "rejeitar" módulos (ramos/raízes) que explorem áreas de solo demasiado secas, sem comprometer o resto do indivíduo. Consequentemente, a arquitetura vascular segmentada incrementa a probabilidade de sobrevivência e retarda a senescência do organismo como um todo, permitindo ainda confinar a entrada de um patogénio ou toxina numa parte restrita: sacrifica-se o módulo para salvar o indivíduo. A setorização hidráulica e proprioceção, claro, não têm um equivalente animal.
Identidade dos módulos
Identidade dos módulos
A identidade dos módulos depende do programa genético expresso a nível meristemático. Um meristema apical caulinar pode começar por produzir módulos com folhas juvenis, em seguida com folhas maduras, depois com brácteas e, finalmente, com as peças da flor. Os novos fitómeros de um caule podem surgir engrossados e diferenciar um tubérculo, ou serem muito curtos e organizarem-se numa roseta de folhas. Muitas plantas anfíbias, como os ranúnculos aquáticos (Ranunculus subgén. Batrachium, Ranunculaceae), transitam abruptamente de programa ontogenético perante a maior ou menor disponibilidade de água livre, exibindo fenótipos alternativos, adaptados a habitats terrestres ou aquáticos (Kim et al., 2018). Geralmente, o câmbio de identidade dos módulos é gradual na conversão das folhas juvenis em adultas e abrupto na diferenciação da flor (v. «Juvenilidade. Indução e diferenciação florais»). A mudança de programa é controlada por uma combinação diversa de fatores endógenos (e.g., determinados pela idade ou a direção dos ramos) ou exógenos (e.g., qualidade da luz, água, fotoperíodo ou temperatura) (Mathews & Kramer, 2012). A identidade, por sua vez, dita a sensibilidade dos módulos ao ambiente; por exemplo, os módulos vegetativos são mais plásticos do que os florais, portanto, o ambiente tem grande influência no desenho do corpo vegetativo, mas menor na estrutura da flor.
No âmbito da biologia do desenvolvimento, a sequência de instruções genéticas expressas a nível meristemático – mais ou menos integradas a diferentes escalas, do raminho até ao indivíduo como um todo –, que orquestra o desenvolvimento de um indivíduo ou de parte dele, constitui o programa ontogenético. O programa ontogenético responde ao ambiente, mostra alguma plasticidade ambiental – esta plasticidade ontogénica [11] [12] , plasticidade desenvolvimental, ontogenic plasticity) é uma componente da plasticidade fenotípica (Charles-Dominique et al., 2012).
Totipotência celular
A natureza séssil das plantas expõe-nas a todo o tipo de danos físicos. Embora a reposição dos módulos perdidos ocorra primariamente a partir de meristemas pré-existentes, estes nem sempre estão disponíveis no local da lesão. Reside aqui a importância da totipotência celular (celular totipotency): a capacidade de uma única célula vegetal de se desdiferenciar (retornar a uma condição meristemática), proliferar e rediferenciar em todos os tipos de células necessários para reconstruir uma planta funcional (Sugimoto et al., 2011). Por conseguinte, a estratégia de construção modular das plantas está intrinsecamente ligada à totipotência celular (Hallé, 2002).
O corpo das plantas contém um vasto número de células somáticas totipotentes no tecido fundamental que enche os seus órgãos (v. «Tecidos definitivos simples»). Quando uma árvore, por qualquer motivo, perde uma fração significativa da sua copa, ativam-se gemas dormentes e/ou diferenciam-se gemas adventícias a partir de células totipotentes, que iniciam a reconstrução da região danificada. Em casos extremos, a copa de uma árvore pode ser totalmente eliminada e em seguida restaurada.
Um exemplo prático é a técnica da rolagem (poda drástica) aplicada aos castanheiros afetados pela doença da tinta (Phytophthora cinnamomi, Heterokontophyta). Ao eliminar a copa, estimula-se a emissão de novos rebentos vigorosos e, consequentemente, a formação de raízes sãs (v. «Setorialização hidráulica»), contendo o avanço da doença no sistema radicular; poucos anos depois, as árvores apresentam uma nova copa e um sistema radicular parcialmente renovado. De igual modo, muitas oliveiras e árvores seculares de parques históricos passaram por vários ciclos de eliminação e reposição da canópia através de ramos epicórmicos (v. «Ramos epicórmicos»). Conclui-se, assim, que a capacidade de reprogramação das células parenquimatosas para uma condição meristemática, conjugada com a estrutura modular, é o segredo da facilidade com que as plantas compensam a perda de órgãos.
Mecanismos análogos envolvendo a totipotência celular explicam a facilidade com que as plantas se propagam vegetativamente, em condições naturais ou pela ação do homem, através de fragmentos de caules, de raízes ou folhas, ou recorrendo a gemas isoladas e aglomerados celulares nas modernas técnicas de micropropagação. Como observou, no final do séc. XIX, o célebre naturalista francês Jean-Henri Fabre: «no que aos animais diz respeito, na imensa maioria dos casos dividir significa destruir; nas plantas dividir é multiplicar» (Mancuso, 2023). Graças a esta capacidade, algumas espécies lenhosas (que regeneram por rebentos de raiz ou de touça) e as plantas herbáceas com intensa propagação clonal (e.g., por afilhamento, rizomas ou bolbos) são virtualmente imortais, uma vez que as partes que colapsam são continuamente substituídas por módulos novos.
Genet e ramet
A arquitetura modular e a frequência da propagação vegetativa na natureza impõem uma distinção terminológica fundamental entre duas categorias de individualidade nas plantas: o genet (indivíduo genético) e o ramet (unidade modular) (Klimešová, 2025). O genet refere-se à totalidade da biomassa com origem num único zigoto, independentemente de as suas partes estarem, ou não, fisicamente conectadas; o ramet constitui a unidade modular potencialmente independente capaz de sobrevivência autónoma se separado da planta-mãe. Esta distinção é crucial em ecologia, uma vez que a contagem de ramets está correlacionada com a densidade e a biomassa, enquanto a contagem de genets avalia a diversidade genética da população.
A maior árvore do mundo em biomassa — e o organismo vivo mais pesado do planeta — é o Pando, um indivíduo clonal de choupo-tremedor (Populus tremuloides, Salicaceae) situado no Utah (EUA). Este gigante estende-se por cerca de 43 hectares e possui uma massa estimada em 6.000 toneladas (Grant et al., 1992). Trata-se de um único genet constituído por um emaranhado de milhares de ramets (neste caso de troncos) que partilham um sistema radicular comum. Fenómenos equivalentes de muito menor dimensão são frequentes em numerosos tipos de ecossistemas. Por exemplo, as manchas de Brachypodium rupestre (Poaceae) e de Urtica dioica (Urticaceae) numa pastagem de montanha, ou um bosquete de Quercus pyrenaica (Fagaceae) no NE de Portugal têm em comum serem, frequentemente, constituídos por um único genet que se espraia em ramets por via clonal.
Numa perspetiva arquitetural, qualquer árvore adulta pode ser entendida conceptualmente de forma análoga: um sistema de ramets — traduzidos nos seus ramos principais ou reiterados — elevados acima do solo por um tronco (Hallé, 2002). Deste modo, a canópia arbórea possui uma natureza intrinsecamente clonal, replicando acima do solo a estratégia de ocupação do espaço que o Pando ou uma gramínea rizomatosa aplicam à superfície do solo.
A enxertia é uma modalidade de propagação vegetativa, omnipresente na agricultura para multiplicar genótipos desejados. Contudo, este processo não é exclusivo da intervenção humana. Sabe-se hoje que a enxertia natural (natural grafting) entre raízes — e, mais raramente, entre ramos aéreos — é frequente em povoamentos de plantas lenhosas coespecíficas ou evolutivamente próximas (Mudge et al., 2009). Esta fusão vascular permite a partilha de água e solutos, criando uma rede cooperativa onde indivíduos genéticos distintos (genets) funcionam como módulos de um sistema maior. Se no caso do Pando um indivíduo atua como uma floresta, aqui a floresta atua como um indivíduo, aumentando a sobrevivência e o sucesso reprodutivo da comunidade (Wohlleben, 2016).
Em todos os exemplos apresentados, estamos perante um mecanismo de integração funcional sem verdadeiro equivalente nos animais superiores.
Modularidade e plasticidade fenotípica
Neste momento será claro para o leitor que a modularidade e a plasticidade fenotípica associada permitem um fino ajustamento da dimensão, da estrutura e da ontogénese (desenvolvimento) das plantas às condições e recursos ambientais – e uma maximização do sucesso reprodutivo (fitness). As plantas têm um corpo flexível em massa, volume e forma. Consoante a sua história de vida, indivíduos coespecíficos com a mesma idade podem ter um tamanho e fisionomias muito distintos. A plasticidade do corpo das plantas não tem paralelo no reino animal. A genética condiciona a plasticidade do fenótipo a dois níveis: i) a maior ou menor capacidade das plantas responderem na forma (e na fisiologia) ao ambiente tem determinantes genéticos: os indivíduos, ecótipos, espécies, etc. não são igualmente plásticos; ii) a resposta plástica varia com o estádio de desenvolvimento; num dado instante é supervisionada pela expressão do programa ontogenético codificado nos genes. O papel da epigenética e os custos da plasticidade fenotípica são discutidos no volume sobre evolução das plantas (v. «Adaptação vs. aclimatação»).
Em resumo, o crescimento por módulos e duas das suas consequências – a cladoptose e a totipotência celular – são essenciais para as plantas ultrapassarem a heterogeneidade ambiental espacial e temporal e as limitações impostas à captura de recursos pela sua natureza séssil (imobilidade). Pela mesma razão, os animais sésseis geralmente também têm uma estrutura modular (e.g., corais). A dormência (v. «Quiescência e dormência dos gomos», «Repouso e crescimento vegetativos das plantas perenes») e outros mecanismos (e.g., ciclo de vida anual) têm uma função similar. Em face da escassez, os animais movem-se em busca de alimento ou morrem; as plantas fazem-se pequenas, deprimem o metabolismo, aquietam-se e, eventualmente, contraem a biomassa viva.
Outras consequências da estrutura modular
A resistência/tolerância à herbivoria das gramíneas pratenses resulta, também, da sua estrutura modular: os animais herbívoros consomem biomassa aérea que posteriormente é restituída por meristemas intercalares e/ou por meristemas axilares (v. «Desenvolvimento e arquitetura das gramíneas»). Em alternativa ou em complemento à reposição, as partes perdidas podem ser compensadas por um crescimento mais vigoroso, mais ou menos descentralizado, de outras(Briske, 1996). Os frutos comerciais têm maior calibre e mais sementes se um número significativo de flores for eliminado com uma poda em verde, por métodos químicos ou por uma geada tardia.
A modularidade e a plasticidade fenotípica a ela associada três outras consequências. (i) Ampliam o nicho ecológico ao permitirem a ocupação de novos habitats sem um período de ajustamento genético de evolução por adaptação – uma das razões por que certas espécies são invasoras bem-sucedidas em novos ambientes distintos dos originais (Levin, 2010). As plantas com flor são mais plásticas do que as restantes plantas vasculares (‘pteridófitos’ + gimnospérmicas), facto que ajuda a explicar o seu sucesso evolutivo (volume II). (ii) Os módulos evoluem de forma quase independente sem alterar significativamente o funcionamento de outras partes. Por exemplo, as flores podem estar sujeitas a uma grande pressão de seleção pelos polinizadores enquanto o corpo vegetativo se mantém inalterado, em estase evolutiva. Nos animais não é assim. Deste modo, a modularidade acelera a adaptação das plantas a novas pressões de seleção, i.e., aumenta a sua capacidade de evoluir (Hansen, 2003). (iii) A competição entre linhagens celulares dentro da própria planta atua como um filtro, eliminando as mutações deletérias antes que estas cheguem às flores/sementes. Por exemplo, uma linhagem de células somáticas sofre uma mutação deletéria, o ramo da planta onde se encontra pode crescer menos vigorosamente e ser suprimido pelos ramos saudáveis. Por conseguinte, a arquitetura modular não é apenas estrutural, mas também uma estratégia evolutiva de "controlo de qualidade" genético, de seleção intra-organismo (ou seleção diplôntica) (Pineda-Krch & Fagerström, 1999).
A natureza modular das plantas torna a sua identificação bastante mais complexa do que a dos animais unitários. Esta será uma das razões por que o número de nomes vernáculos tradicionais (não trabalhados pelos taxonomistas como acontece no inglês), cujo conceito coincide com os taxa definidos pelos taxonomistas, é geralmente limitado no mundo das plantas. Muitos animais podem ser positivamente identificados com base na silhueta, no tamanho, na cor, na postura corporal ou até no movimento. Por isso, as aves ou os mamíferos são representados em corpo inteiro nos guias de campo. A fisionomia das plantas é francamente menos informativa. A identificação das plantas obriga a uma observação visual de proximidade, por vezes com recurso a lupas de bolso, da inserção e forma das folhas, dos pelos das folhas e do cálice, da estrutura da flor, dos frutos, entre outros aspetos. Ainda assim, sobretudo em ambientes tropicais, a forma das árvores e arbustos pode ser de grande utilidade para determinar a identidade de indivíduos. Quando se observam povoamentos arbóreos em fotografia aérea, ou em contraluz, é relativamente fácil distinguir a silhueta de um castanheiro, de uma tília ou de um pinheiro-manso.
Simetria
A simetria (simetry), por definição, consiste na repetição regular, geneticamente determinada, de elementos estruturais iguais ou similares (Figura I.1.11). A maioria dos animais, mais concretamente os taxa do clado dos Bilateria (inc. artrópodes e vertebrados), tem uma simetria bilateral dorsiventral definida nas primeiras fases da embriogénese – que emerge diretamente da direcionalidade do seu movimento sob o efeito da força da gravidade (Westerkamp & Classen-Bockhoff, 2007). As patas dos insetos estão sempre dirigidas para a interface com o substrato numa posição ventral. As asas, necessariamente, estão inseridas por cima das patas. As simetrias externas das plantas são bem mais complexas (Quadro I.1.4).

FIGURA I.1.11. Simetria. A) Simetria radial: flor de Hibiscus rosa-sinensis (Malvaceae); n.b.: estames monadelfos. B) Simetria bilateral: folhas dorsiventrais de Quercus lusitanica (Fagaceae). C) Simetria por metameria: caule de amor-de-hortelão (Galium aparine, Rubiaceae). [Fotografias do autor.]
|
QUADRO I.1.4 TIPOS DE SIMETRIA |
||
|
Tipo de simetria |
Descrição |
Exemplo |
|
Simetria por metameria |
Repetição de elementos estruturais ao longo de um eixo. |
Repetição do módulo elementar caulinar – o fitómero – constituído por um entrenó, um nó e uma ou mais folhas com os respetivos meristemas axilares. |
|
Simetria radial |
Repetição de um número variável de elementos estruturais, com um mesmo ângulo, em torno de um eixo. |
Pétalas em redor do eixo floral (= recetáculo) ou a disposição radial dos tecidos no caule e na raiz. |
|
Simetria bilateral |
Repetição de elementos estruturais nos dois lados de um plano de simetria. Simetria predominante no reino animal. |
Flores zigomórficas (com um plano de simetria) e folhas dorsiventrais. Comum nas flores polinizadas por insetos. |
As regras das simetrias por metameria e radial codificadas no genoma controlam a disposição espacial dos módulos elementares que constituem o corpo das plantas (fitómeros). A forma dos indivíduos resulta da interação dessas regras com o ambiente – e.g., com a disponibilidade de luz, água e nutrientes no solo –, existindo, para tal, sensores especializados das características ambientais (e.g., fitocromo). Uma programação completa do corpo das plantas, além de incompatível com a volatilidade temporal intrínseca dos habitats das plantas e com os riscos criados pela imobilidade, exigiria muito mais informação genética do que a requerida na programação da metameria, das simetrias radial e bilateral, da estrutura dos fitómeros, dos sistemas fisiológicos de sensores e da integração da informação sensorial. As regras de simetria, conjugadas com a estrutura modular, são, então, uma solução evolutiva parcimoniosa (simples e que exige pouca informação) para gerar, em resposta ao ambiente, formas complexas e plásticas, mas, ao mesmo tempo, evolutivamente flexíveis, i.e., sensíveis a pressões de seleção de índole diversa. A prontidão para mudar a estrutura e a fisiologia do corpo é uma característica fundamental das plantas.
Crescimento indeterminado nas plantas
Numa fase precoce do desenvolvimento embrionário das plantas, a produção de novas células passa a estar concentrada em tecidos especializados designados por meristemas. Nos animais, pelo contrário, a divisão celular ocorre de forma difusa, com diferentes atividades mitóticas um pouco por todo o corpo, sendo mais intensas antes de atingida a fase adulta. As células meristemáticas das plantas são funcionalmente análogas às células estaminais animais, no sentido em que são histogénicas, i.e., diferenciam tecidos especializados. As plantas diferem ainda dos animais na capacidade adicional dos meristemas apicais gerarem órgãos vegetativos (e.g., raiz, caule e folhas) ou reprodutivos completos (e.g., anterídios, cones e flores).
As plantas estão «condenadas» a crescer, ainda que com interrupções cíclicas, até à senescência ou morte. Uma planta pode ter centenas de anos, mas é anualmente reabastecida de tecidos jovens provenientes dos meristemas. A sua imobilidade confere-lhes uma grande suscetibilidade a pressões externas, destacando-se seis causas principais para a necessidade deste crescimento indeterminado: (i) a herbivoria (fitofagia), (ii) o parasitismo, (iii) a competição pela luz, (iv) a depleção dos nutrientes no solo, e a degradação dos sistemas (v) fotossintético e (vi) vascular.
A produção de esporos (nas ‘plantas de esporulação livre’), sementes (nos espermatófitos) ou propágulos constituem os únicos momentos do ciclo de vida em que as plantas dispõem de oportunidade para se «desenraizarem» e escapar a ameaças existenciais externas. Contudo, estes são eventos demasiado raros e insuficientes, se se tiver em consideração o rácio entre a duração da dispersão e a extensão do ciclo fenológico.
A reposição pelo crescimento de partes perdidas é a grande resposta dos vegetais aos desafios criados pela herbivoria e pelo parasitismo. O movimento do fósforo e dos nutrientes catiónicos no solo é limitado; uma vez esgotada a reserva local, a solução é partir em busca de solo não explorado, construindo mais raiz. O acesso à luz depende da emissão de caules com folhas acima da canópia dos competidores diretos. A degradação dos sistemas fotossintético e vascular com o tempo é, igualmente, resolvida pela continuidade do crescimento, uma vez que a capacidade de reparação destas funções à escala celular é limitada. As folhas «danificadas» são substituídas por novas folhas e o câmbio vascular produz novo tecido condutor. Em resumo, nas plantas, o funcionamento do corpo é assegurado pela adição, reposição ou substituição de partes, ao nível do tecido ou do órgão. Nos animais, pelo contrário, a manutenção ocorre essencialmente a nível celular – a substituição e a reparação de tecidos e órgãos inteiros são, consoante os casos, limitadas ou inexistentes. Ao contrário dos animais, nas plantas, parar de crescer é morrer.
A produção de novas folhas implica, mais tarde ou mais cedo, a eliminação das folhas mais velhas disfuncionais. O turnover das raízes finas é também muito elevado (v. «Crescimento e Arquitetura do sistema radicular»). Parte dos caules é eliminada por cladoptose (v. «Cladoptose»). Ainda assim, as plantas retêm uma vasta massa de partes não vivas com função de suporte, concretamente xilema e tecidos associados, nas camadas mais profundas do seu corpo (v. «Xilema secundário»). Este mecanismo de acumulação, pelo menos com esta dimensão, não tem paralelo nos animais.
As estruturas reprodutivas das plantas, ao invés do corpo vegetativo, têm uma organização hierárquica e uma estrutura determinada. Na flor completa, as pétalas sucedem-se às sépalas, os estames às pétalas e o gineceu ao androceu. Esta sequência tem um controlo genético preciso. Com a formação da flor, extingue-se o meristema que lhe deu origem. A dimensão e a estrutura das partes dos órgãos reprodutivos das plantas são menos plásticas do que o corpo vegetativo. Este facto, somado à diversidade morfológica e funcional das estruturas reprodutivas, explica a importância da flor na identificação e classificação das plantas terrestres.
Mecanismos de degradação dos sistemas fotossintético e vascular
Tendo em consideração a sua importância funcional, sem nada que se lhe pareça entre os animais, convém explorar um pouco melhor os mecanismos de degradação dos sistemas fotossintético e vascular. Os sistemas vasculares e fotossintético das plantas são frágeis e temporariamente eficientes. Os mecanismos envolvidos na sua degradação estão bem esclarecidos.
A água que preenche os vasos e traqueídos do xilema encontra-se sob tensão. Nestas condições, os gases têm tendência a libertar-se da solução formando bolhas microscópicas que podem coalescer, formar bolhas maiores, e obstruir os elementos traqueais do xilema e romper as conexões xilémicas nos caules e raízes. Nos climas frios, os ciclos de congelação-descongelação têm o mesmo efeito. O processo de formação de bolhas gasosas designa-se cavitação (cavitation); a cavitação precede e causa a embolia (embolism) do xilema. As plantas têm mecanismos para conterem a propagação da embolia (e.g. formação de tiloses e sistema tórus-margo) que não evitam, porém, uma lenta perda de eficiência do sistema condutor (i.e., da condutância hidráulica). A embolia do xilema está relacionada com a lenta recuperação ou a morte parcial ou total das plantas sob stresse hídrico. A resistência do xilema à embolia é adaptativa e chave na estruturação das comunidades vegetais em função da disponibilidade de água no solo (Blackman et al., 2014).
A capacidade de conversão da energia da luz em cadeias carbonadas de alta densidade energética vai decaindo à medida que as folhas vão envelhecendo. Por quatro razões:
- Fotodegradação do sistema fotossintético – a incidência da luz nas folhas induz a formação de radicais livres que danificam as membranas cloroplásticas e os fotopigmentos; a eficiência fotossintética[13] começa a decrescer logo após a expansão das folhas, muito antes destas entrarem em senescência;
- Degradação mecânica, herbivoria e parasitismo das folhas – as folhas estão sujeitas a abrasão (e.g., danos causados por grãos de areia ou sais transportados a grande velocidade pelo vento) e rasgam-se, sendo consumidas ou parasitadas com alguma facilidade;
- Ensombramento das folhas por efeito do crescimento – uma vez que os ramos, enquanto funcionais, se alongam continuamente, as folhas mais velhas vão ficando relegadas para as camadas mais profundas da copa, cada vez mais ensombradas, até que os seus consumos respiratórios de energia ultrapassam os ganhos fotossintéticos;
- Acumulação de substâncias tóxicas – as folhas degradam-se naturalmente pela acumulação de substâncias tóxicas transportadas dissolvidas na corrente transpiratória a partir do solo, ou produzidas pelo metabolismo secundário..
Sexualidade e ciclo de vida
No estudo das plantas-terrestres, considera-se que o ciclo de vida (= ciclo biológico de vida) se estende da germinação do esporo (nas ‘plantas de esporulação livre’) ou da semente (nos espermatófitos) – eventualmente, desde o início da atividade de estruturas de multiplicação vegetativa (ex. estolhos e rizomas) – até à dispersão ou senescência (morte). No capítulo IV «Ciclos floral e reprodutivo das angiospérmicas» será usado um conceito distinto. Ciclo de vida não é o mesmo que ciclo floral ou de ciclo reprodutivo (v. «Ciclos floral e reprodutivo das angiospérmicas»).
O ciclo de vida é diplonte nos animais, e haplonte (tipo ancestral característico de muitos grupos algais) ou haplodiplonte nas plantas (v. «Ciclos de vida das ‘plantas de esporulação livre’»). Os animais possuem um único tipo de célula reprodutiva: o gâmeta. O ciclo haplodiplonte característico das plantas terrestres envolve a alternância de dois tipos, morfologicamente distintos, de corpo multicelular – o esporófito e o gametófito –, especializados na produção de distintas células reprodutivas unissexuais, respetivamente, esporos e gâmetas. O gametófito e o esporófito têm histórias evolutivas distintas.
Num estádio inicial do ciclo de vida dos animais diferencia-se um grupo especializado de células – as células germinais (germ cells) – do qual derivarão os gâmetas. Este mecanismo é designado por Barreira de Weismann. Por conseguinte, nos animais as mutações somáticas (em células não reprodutivas) não são transmitidas à descendência. Nas plantas terrestres, as células precursoras dos gâmetas não estão predeterminadas no embrião – o destino das células produzidas nos meristemas é flexível (Hallé, 2002). Consequentemente, as plantas tendem a acumular mais mutações somáticas do que os animais porque os gâmetas são produzidos a partir de linhagens de células somáticas que sofreram múltiplas divisões e acumularam erros de cópia – e nada impede que uma mutação ocorrida num meristema situado num determinado ponto da copa possa ser transmitida, por via assexual ou sexual, à descendência (Lanfear, 2018; Schoen & Schultz, 2019). A modularidade dos vegetais permite que a própria experiência somática do indivíduo contribua diretamente para a diversidade genética da população.
As células da canópia das plantas com semente estão expostas a doses elevadas de radiação UV. As radiações mutagénicas incrementam a variação genética e aceleram as taxas de evolução, em particular nas espécies em que prepondera a reprodução assexuada. Em contrapartida, representam um risco de envelhecimento genético (acumulação de mutações somáticas deletérias) precoce, sobretudo nas plantas mais longevas como as grandes árvores. Quanto mais tempo vive uma planta, mais prolongada a exposição à radiação, maior o número de divisões celulares (a maior parte das mutações ocorre durante a mitose), e maior a probabilidade de se acumularem mutações somáticas deletérias. Os catafilos ou os esboços foliares, que envolvem as células meristemáticas apicais e axilares caulinares, e a formação de um ritidoma por fora do câmbio contêm as radiações mutagénicas, mas não chegam. O envelhecimento genético nas plantas é mitigado por dois mecanismos: (i) meiose e (ii) células iniciais. A meiose, o mais importante dos dois, é comum a plantas e animais (mecanismo discutido no volume II). A taxa mitótica das células meristemáticas não é constante: as células ditas iniciais dividem-se raramente dando origem a outras células, essas sim que proliferam em grande número (v. «Tipos celulares meristemáticos»). Portanto, a ocorrência de um grupo de células iniciais reduz a acumulação de mutações deletérias, efeito confirmado experimentalmente em grandes árvores seculares (Schmid-Siegert et al., 2017). As células iniciais nas plantas e as células germinais nos animais são duas soluções para um mesmo problema: o envelhecimento genético.
Os animais movem-se, os sexos encontram-se; nos grupos mais avançados, a escolha dos melhores parceiros, e, implicitamente, dos melhores genes, envolve complexos rituais e uma pormenorizada avaliação do fenótipo (seleção sexual); por fim, realiza-se a transferência de gâmetas – o ato sexual. A imobilidade das plantas fá-las, em grande medida, dependentes de vetores (e.g., insetos, vento ou a água) para o transporte de pólen, i.e., dos gâmetas masculinos. A seu tempo veremos que a flor é entendida como uma adaptação à polinização (v. «Polinização»). A seleção dos melhores genes faz-se através da competição do pólen ao nível do estigma ou do pistilo (v. «Competição do pólen. Seleção de gâmetas») (Skogsmyr & Lankinen, 2002).
A embriogénese das plantas terrestres (embriófitos) desemboca numa estrutura simples – o embrião –, um rudimento do esporófito que nas plantas com flor se resume a um meristema apical radicular, uma raiz embrionária, o hipocótilo, um ou dois cotilédones, o epicótilo e a plúmula que contém o meristema apical caulinar (Figura 1.4-B). Ao contrário da maioria dos animais, o embrião pode permanecer dormente um longo período de tempo. Todos os órgãos das plantas adultas são diferenciados durante a vida pós-embrionária por meristemas que produzem órgãos completos (raiz, caules, folhas e flores). As células, os tecidos e os órgãos das plantas adultas, ao contrário do ocorrido nos animais unitários, não têm a mesma idade. As células diminuem de idade de dentro para fora nos caules com crescimento secundário (Figura 81); as folhas e os tecidos caulinares proximais são mais velhos do que os distais.
[1] Folhear ou foliar.
[2] Designação do cotilédone nas poáceas.
[3] A monofilia das arqueplastidas não é recuperada com elevados níveis de certeza em muitos estudos filogenéticos recentes (Burki et al., 2020). Esta incerteza desafia o dogma da endossimbiose primária única, sugerindo cenários alternativos complexos, como múltiplos eventos de endossimbiose ou a transferência lateral de plastídeos entre linhagens ancestrais.
[4] As plantas parasitas e as plantas saprófitas são secundariamente heterotróficas. Secundariamente porque os ancestrais das plantas heterotróficas eram clorofilados e fotoautotróficos.
[5] De metamorfoses completas (e.g, himenópteros, coleópteros e lepidópteros); opõe‑se à hemimetabolia (e.g., homópteros).
[6] A totipotência celular é a capacidade de uma célula retomar a capacidade de se multiplicar (= capacidade meristemática) e dar origem a todos os tipos de células diferenciadas do organismo.
[7] Na bibliografia, discute‑se se o entrenó do fitómero é anterior ou posterior ao nó. Na prática, esta decisão é irrelevante.
[8] As flores são o módulo reprodutivo das angiospérmicas, por sua vez decomponível em módulos elementares (fitómeros) constituídos pelo verticilo floral mais o curto entrenó de recetáculo que o sucede.
[9] As características mencionadas para o trevo-branco podem, porem, ser fixadas por seleção. Estão em causa dois processos distintos: plasticidade fenotípica vs. adaptação. Uma e outra são destrinçáveis através da transplantação para ambientes controlados: a plasticidade fenotípica é reversível, a adaptação não (embora os genótipos adaptados à herbivoria possam sofrer algumas modificações fruto da sua plasticidade fenotípica intrínseca).
[10] Um exemplo clássico de plasticidade fenotípica: a dimensão dos entrenós e das folhas é influenciada pela exposição à luz: os ramos mais expostos ao sol têm, frequentemente, folhas mais pequenas, enquanto os ramos estiolados exibem entrenós mais longos e folhas maiores e mais delgadas, de cutícula menos espessa, e ricas em clorofila.
[11] O conceito de plasticidade ontogénica pode ser aplicado a órgãos, módulos, partes das plantas, ao sistema radicular, à canópia ou à planta como um todo.
[12] Importante não confundir os dois tipos fundamentais de plasticidade referidos neste livro: evolutiva, fenotípica (inc. pasticidade ontogénica).
[13] Fração da energia radiante incidente que é convertida em energia química (fotoassimilados). Consoante os propósitos da investigação, a eficiência fotossintética pode: referir‑se à radiação solar global ou apenas à radiação fotossinteticamente ativa (PAR); ser quantificada através do CO2 fixado, do O2 libertado ou da biomassa acumulada (e.g., por indivíduo ou por unidade de superfície); e reportar-se a uma medição instantânea ou integrada num determinado período de tempo.